Зачем нашему организму нужны антиоксиданты?
В наше время мы все чаще слышим термин «антиоксидантные свойства», но зачастую не знаем достоверно, что же означает это выражение. Давайте разберемся как влияют на организм антиоксиданты и какие продукты содержат наибольшее их количество.
Основная задача антиоксидантов — нейтрализовать окислительные процессы в организме, которые оказывают разрушительное действие на наш организм. Чтобы детальнее разобраться в природе антиоксидантов, давайте сначала рассмотрим действие их противоположностей – оксидантов. Оксиданты – вещества, которые появляются в нашем организме в результате нарушения обменных процессов, которые являются следствием неправильного питания, стрессов, плохой экологии и т.д. С химической точки зрения, оксидант – это элемент, у которого есть неспаренные электроны. Недостающие электроны оксидант компенсирует за счет здоровых клеток, таким образом окисляя их и разрушая их структуру. Такие процессы очень опасны для организма и могут привести к появлению онкологических заболеваний.
Антиоксиданты, как уже упоминалось выше, нейтрализуют процесс окисления. Происходит это за счет снабжения оксидантов недостающими электронами и как следствие – восстанавливается химическая связь в клетках организма.
Антиоксидантные свойства наиболее выражены в витаминах С и Е, бета-каротине, ликопине, танине, антоциане. Все эти вещества наш организм получает из пищи.
Витамин С
Самый известный из всех витаминов и один из самых мощных антиоксидантов. У всех витамин С ассоциируется, в первую очередь с цитрусовыми. Но это не единственный его источник. Большое количество этого вещества содержится в ягодах (особенно в черной смородине и шиповнике), шпинате, брюссельской капусте.
Витамин Е
Витамин Е кроме устранения окислительных процессов также способствует усвоению витамина А. Большая концентрация витамина Е есть в орехах, семенах, стручковой фасоли, горохе, в растительных маслах и рыбьем жире.
Витамин А
Этот витамин также принимает активное участие в восстановительных процессах и способствует нормализации клеточного обмена. Основные источники витамина А: морковь, брокколи, тыква, персики, абрикосы, птрушка.
Основные источники витамина А: морковь, брокколи, тыква, персики, абрикосы, птрушка.
Ликопин
Ликопин – вещество, благодаря которому некоторые фрукты и овощи имеют ярко выраженный красный цвет. Этот элемент не может синтезироваться организмом и постуает исключительно с пищей. Наибольшее количество ликопина содержится в томатах, красном болгарском перце, грейпфрутах, а также в большинстве овощей и фруктов красного цвета.
Танины и антоцианы
Танины – это вещества естественного происхождения, которые содержатся в основном в семенах, листьях и кожуре растений и придают им терпкий вкус. Антоцианы – пигментные вещества, которые также находятся в растениях и придают им красный, фиолетовый и синий цвета. Танины содержатся в хурме, зеленом чае, какао, шоколаде, красном вине. Антоцианами богаты черника, ежевика, вишня, темный виноград, баклажаны.
Следует отметить, что на сегодняшний день действие антиоксидантов еще детально изучается, но уже сейчас можно с уверенностью сказать, что антиоксиданты – это та сила, которая помогает организму противостоять различным заболеваниям и разрушительным процессам.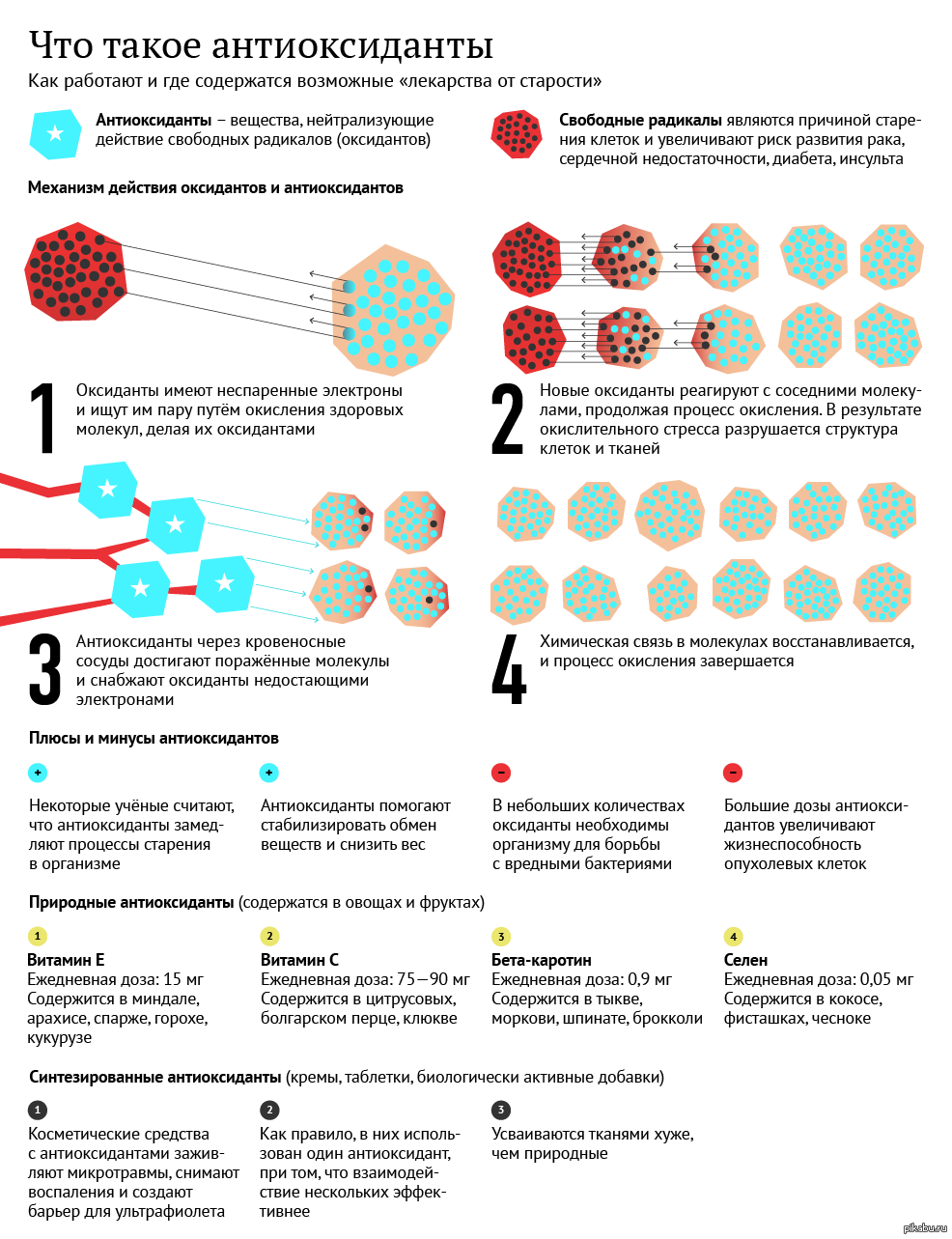
Антиоксиданты для укрепления иммунитета
Статья о роли антиоксидантов в нашем организме, а также о том как они влияют на иммунную систему, что особенно актуально сейчас.
СодержаниеЧто такое антиоксиданты
Антиоксидант дословно означает «противодействующий окислению».Каждый день наши клетки сталкиваются с действием свободных радикалов — оксидантов, в составе которых не хватает одного или нескольких электронов, чтобы стабилизироваться, отбирают необходимые электроны у здоровых клеток и тканей. Это и называется окислением, которое в результате приводит к старению, снижению иммунитета, а также возможно разрушение клеток и тяжелые заболевания.
Свободные радикалы попадают в организм, а также вырабатываются самим организмам в результате плохой экологии, неправильного питания, вредных привычек и неправильного образа жизни, а также с ультрафиолетом и радиацией.
Свободные радикалы в большом количестве могут приводить к серьезным болезням:
Вряд ли в современном мире нам удастся избежать попадания свободных радикалов в организм, однако, использование антиоксидантов позволит защитить клетки нашего организма от вредного действия оксидантов.1. Болезни Альцгеймера и Паркинсона.
2. Онкологические заболевания.
3. Варикозная болезнь, атеросклероз сосудов, тромбоз.
4. Бронхиальной астмы.
5. Сахарный диабет.
 Болезни Альцгеймера и Паркинсона.
Болезни Альцгеймера и Паркинсона.Как действуют антиоксиданты
Как я уже писал выше, свободные радикалы забирают из клеток электрон, так вот у антиоксидантов есть лишние электроны, которые позволяют нейтрализовать свободные радикалы, либо восстановить поврежденные клетки.Какие существуют антиоксиданты
Натуральными антиоксидантами являются:
- витамин А
- витамин С
- витамин Е
- селен
- цинк
- дигидрокверцетин
- карнозин
- полифенолы
- катехины

Антиоксидантами богаты: миндаль, арахис, грецкий орех, подсолнечное масло, авокадо, спаржа, горох, кукуруза, цитрусовые, кислые ягоды (смородина, клюква), зелень (петрушка, брокколи, шпинат), тыква, морковь, абрикосы, персики, манго, рыба, морепродукты, чеснок, фисташки.
Роль антиоксидантов в иммунной системе.
Обычно организм контролирует количество свободных радикалов, но в период вирусов и инфекций, возбудители заболеваний попадают в организм в большом количестве. Собственные антиоксиданты быстро расходуются и не успевают синтезироваться, поэтому число свободных радикалов резко растет. Кроме того, «запасы» антиоксидантов истощаются при курении, приеме алкоголя, а также при хронических заболеваниях.- Разрушают клетки иммунной системы, не позволяя им выполнять свои функции, и тем самым ослабляют иммунитет.
- Повреждают пораженные вирусами или бактериями ткани, и поэтому усиливают воспаление. А воспаление, в свою очередь, вызывает образование новых свободных радикалов.

Следовательно, чтобы дать своему организму возможность эффективно бороться с инфекцией, следует обеспечить его антиоксидантами разного механизма действия.
Какие антиоксиданты выбрать?
Большинство людей знают популярные витамины, обладающие антиоксидантным действием, также есть значительное количество растительных экстрактов оказывающих антиоксидантный эффект:Витамин С – самый известный витамин и антиоксидант. Нейтрализует свободные радикалы, а также оказывает противовоспалительное действие и служит катализатором большого количества важных физиологических процессов.
Витамин Е нейтрализует сразу несколько видов свободных радикалов и препятствует их проникновению в клетку, также защищает клеточные мембраны от повреждения свободными радикалами.

Витамины А, С, Е укрепляют кровеносные сосуды и предохраняют их от повреждения, снижают вязкость крови, восстанавливают микроциркуляцию, снижают тромбообразование.
Селен является «ловушкой» свободных радикалов и обрывает реакции окисления. Селен положительно влияет на процессы синтеза в клетках: увеличивает синтез ДНК, РНК, белка, гемоглобина, АТФ, улучшает качество жизнедеятельности мужских половых клеток. Селен позитивно влияет на иммунную систему.
L-карнозин защищает организм от закисления, благодаря способности связывать ионы водорода, а также нормализует психо-эмоциональное состояние, снижает стресс, улучшает сон.
Цинк является активным центром важного фермента — супероксиддисмутазы, который обеспечивает антиоксидантную защиту организма. Цинк участвует в синтезе разных анаболических гормонов в организме, включая инсулин, тестостерон и гормон роста.

Желательно принимать не один антиоксидант, а их комплекс, так как вместе они действуют намного лучше и часто оказывают взаимоусиливающий эффект.
Автор: Адам Хасанов подробнее
Промокод: article введите данный промокод при оформлении заказа
в нашем интернет-магазине и получите скидку 20% на весь заказ!
Теорию старения придется переписать — Российская газета
Новые научные данные ставят под сомнение долгие годы считавшуюся бесспорной теорию старения. Главный виновник многих болезней и раннего старения наукой давно назван. Это так называемые свободные радикалы (оксиданты), которые повреждают клетки, запуская в организме различные разрушительные процессы.
Кстати, радиация, гербициды, различные поражающие воздействия вызывают в организме настоящий взрыв свободных радикалов. Вычислив врага, наука нашла и защиту от него — антиоксиданты. Ученые утверждают, что антиоксиданты блокируют действие свободных радикалов, уменьшают их вредное действие. Отсюда как раз повальное увлечение БАДами, витаминами и диетами.
Этой теории почти 50 лет, по сути, она стала краеугольным камнем геронтологии. Но в последние пять лет ученые стали получать неожиданные данные. Так, британский исследователь из Лондонского университета Дэвид Джемс у одного из видов червей отключил гены, ответственные за выработку антиоксидантов. Он предполагал, что, как и положено по теории, свободные радикалы, не встречая сопротивления, расцветут буйным цветом, и черви быстро умрут. Однако они и не собирались это делать.
Но это было только начало. Американский исследователь из Техасского университета Рошель Баффенштайн пыталась выяснить, почему рекордсменом по продолжительности жизни среди грызунов являются слепыши: они живут в восемь раз дольше.
Но самый сильный удар по теории оксидантов нанес Зигфрид Хекими из Канадского университета Макгилла. Он вывел генетически модифицированный вид червей, у которых активно вырабатывались свободные радикалы. Подопытные жили на 30 процентов дольше обычного. Зато, когда им стали скармливать антиоксидант витамин С, эффект долгожительства исчез.
И таких экспериментов, которые ставят под сомнение оксидантную теорию старения, становится все больше. Означает ли это, что ее надо закрывать и браться за разработку новой? Однозначного ответа пока нет. На данный момент ученые считают, что в одних ситуациях свободные радикалы могут быть вредны, а в других, наоборот, полезны. Скажем, в очень больших количествах они действительно вызывают множество повреждений в клетках и могут стать причиной рака. Зато при небольшом содержании они, напротив, вместо ускорения старения его замедляют.
Еще недавно витамины, разные пищевые добавки считались безусловным благом, но теперь это поставлено под сомнение. Более того, выявлено, что у тех, кто принимает бета-каротин и витамины А и Е, выше риск различных заболеваний, чем у не принимавших никаких витаминов. Ученые в растерянности. Пока они не могут объяснить сенсационные эксперименты. Но одно ясно: теорию старения придется кардинально переписать.
Оксиданты и антиоксиданты — ПИСЬМА АЛЕКСУ — LiveJournal
То, что я собираюсь сказать, возможно, покажется ересью с точки зрения современной науки о здоровье.
Считается, что окисление (оксидация) — это плохо, а то, что препятствует окислению — антиоксиданты — это хорошо.
Скажу только, что заинтересованные лица и компании делают на этом представлении очень хорошие деньги.
Антиоксиданты полезны для вашего здоровья, и поэтому должны стоить дороже. Например, кофе — это оксидант, а чай, особенно зеленый — антиоксидант. Хотя наиболее активное действующее вещество у обоих — это кофеин. Как такое может быть?
Хотя наиболее активное действующее вещество у обоих — это кофеин. Как такое может быть?
Сахар — тоже оксидант, и многие платят этому серьезную дань. Диабет, и т.п. окислительные приключения.
Давайте вспомним, какие процессы имеют в своей основе окисление. Горение, любой огонь, дающий тепло — это окисление (оксидация).
Насыщение эритроцитов кислородом в легких — это окисление. Элемент железо в крови ржавеет (красные эритроциты) — это окисление. Fe становится Fe2O3, оксид железа, это то, чего вы так не любите на вашем клинке, да и просто на вашей Тойоте Камри, но это то, что делает вас живым. Вы стараетесь счистить ржавчину с вашего меча — это делает его острее. Вы стараетесь счистить ржавчину с ваших эритроцитов — это делает вашу жизнь тупее.
Табак, алкоголь, наркотики и их воздействие на кровь — это окисление, сокращающее жизнь, но делающее ее интенсивнее. Секс, оргазм — это реакция окисления, сокращающая жизнь. А вы как думали?!
Все это о том, как долго вы хотели бы жить? Но это также и о том, как, насколько интенсивно вы хотели бы жить?
Если вы хотите жить долго — вы выбираете антиоксиданты.
Если вы хотите жить интенсивно и насыщенно — вы выбираете оксиданты.
Если вы мудры — вы выбираете баланс между этими двумя.
Все больше уважаю Макаревича.
В начале — его песня о выборе между оксидантами и антиоксидантами.
PS: Важно еще ЧТО, какие начала в мире вы питаете своей энергией и греете своим огнем, если выбираете гореть? И, если вы выбираете антиоксиданты, ради ЧЕГО вы пытаетесь сохранить энергию и жить долго?
Обратите внимание также на слова «И Бог хранит меня». Это то, чего многие не учитывают, когда выбирают между оксидантами и антиоксидантами — вмешателство высших энергий, имеющих свои интересы в том, что происходт на Земле.
Антиоксиданты — AhmadTea
14 Фев, 2020
Категории: Новости История Мир чая
Антиоксиданты, буквально «против-окислители», вещества, которые, попадая в организм человека, находят и нейтрализуют врагов здоровых клеток — свободные радикалы.
Сводные радикалы — это продукт распада клетки, поврежденные молекулы, появлению которых в нашем теле способствует плохая экологическая ситуация и дурные привычки, в особенности — курение и алкоголь. Свободные радикалы «выбирают» из здоровой клетки протеины и жиры, нарушая структуру ДНК и даже убивая здоровую клетку. Это, в свою очередь, способствует старению организма, развитию различных заболеваний, в том числе онкологических и сердечно-сосудистых.
Свободные радикалы «выбирают» из здоровой клетки протеины и жиры, нарушая структуру ДНК и даже убивая здоровую клетку. Это, в свою очередь, способствует старению организма, развитию различных заболеваний, в том числе онкологических и сердечно-сосудистых.
Антиоксиданты — природные помощники человека в борьбе с многочисленными напастями. Самые известные антиоксиданты — витамины С и Е, бетакаротин, селен, биофлавонаиды. Их число особенно велико во фруктах, овощах, злаках, красном вине и чае, особенно зеленом. Именно зеленый чай считается самым богатым кладезем антиоксидантов. В зеленом чае антиоксидантов больше, чем в черном где-то в десять раз.
При регулярном потребление зеленого чая (4-6 чашек в день) его антиоксиданты оказывают значительное укрепляющее и профилактическое действие на весь организм, понижают уровень разрушения клеток и даже способствуют их восстановлению.
Антиоксиданты снижают уровень «плохого» холестерина в крови, тормозят процесс отложения «бляшек» на стенках кровеносных сосудов. Они также имеют свойство растворят тромбы, предотвращают их образование и способствуют разжижению крови, что особенно важно для людей преклонного возраста.
Они также имеют свойство растворят тромбы, предотвращают их образование и способствуют разжижению крови, что особенно важно для людей преклонного возраста.
Еще один «плюс» антиоксидантов — способность сохранять здоровье десен и зубов, так как они успешно борются с бактериями, населяющими полость рта. Антиоксиданты являются прекрасным природным лекарством против кариеса. К антиоксидантам зеленого чая относятся так называемые полифенолы, а конкретно — четыре вида катехинов и кверцетин. Они сохраняют свою активность в организме человека примерно в течение двух часов. Необходимо отметить, что чай с лимоном продлевает деятельность антиоксидантов, а добавление в чай молока не оказывает на них никакого пагубного воздействия.
Зеленый чай — это эликсир молодости и здоровья, природное «средство Макропулоса», которое реально может продлить жизнь при ежедневном употреблении. Особенно вкусны и полезны высококачественные чаи из коллекции Ahmad Tea, например, Жемчужина Королевского Дракона или Сенча Кабусэ.
Что могут антиоксиданты
Похоже, наш организм настолько мудр, что не нуждается в дополнительных антиоксидантах?
С. Е.: Как раз наоборот – некоторые из них мы можем получить только извне, например, витамины Е, С, флавоноиды, ликопин. Растительная пища – самый надежный и естественный источник полезных веществ с антиоксидантными свойствами.
Как антиоксиданты влияют на процесс старения?
С. Е.: Активные формы кислорода (оксиданты извне или изнутри) воздействуют на генетический аппарат клеток и вызывают их естественную смерть (апостоз). Тем самым они ускоряют процесс старения, а антиоксиданты (теоретически) могут его замедлить – этому есть экспериментальные подтверждения. Однако добиться полного, научно обоснованного контроля над старением ученым пока не удается. Нам еще предстоит понять, как, в каких дозах и какие именно антиоксиданты нужно для этого использовать.
Насколько эффективны эти вещества в косметике?
С. Е.: В антивозрастных средствах используются вещества, действие которых хорошо изучено. Например, витамины С и Е или метаболитный антиоксидант под названием Q10, который вырабатывается в нашем организме, но с возрастом становится дефицитным. Их эффективность связана с тем, насколько легко эти антиоксиданты проникают в кожу. Скажем, жирорастворимый витамин Е хорошо усваивается кожей. А водорастворимые витамин А и бета-каротин проникают только в ее поверхностные слои. Если антиоксиданты «упакованы» в нанокапсулы (микрочастицы, размер которых позволяет транспортировать заключенные в них вещества в глубь тканей, не повреждая их), они, как правило, эффективны – но далеко не все косметические марки могут позволить себе эту дорогостоящую технологию. Однако стоит иметь в виду, что вид нашей кожи в большей степени зависит от того, насколько эффективно она питается изнутри, через кровеносную систему.
Е.: В антивозрастных средствах используются вещества, действие которых хорошо изучено. Например, витамины С и Е или метаболитный антиоксидант под названием Q10, который вырабатывается в нашем организме, но с возрастом становится дефицитным. Их эффективность связана с тем, насколько легко эти антиоксиданты проникают в кожу. Скажем, жирорастворимый витамин Е хорошо усваивается кожей. А водорастворимые витамин А и бета-каротин проникают только в ее поверхностные слои. Если антиоксиданты «упакованы» в нанокапсулы (микрочастицы, размер которых позволяет транспортировать заключенные в них вещества в глубь тканей, не повреждая их), они, как правило, эффективны – но далеко не все косметические марки могут позволить себе эту дорогостоящую технологию. Однако стоит иметь в виду, что вид нашей кожи в большей степени зависит от того, насколько эффективно она питается изнутри, через кровеносную систему.
Так как же нам себе помочь?
С. Е.: Наиболее эффективны антиоксиданты в натуральных продуктах, поскольку они обладают высокой биодоступностью, то есть легко проникают в кровь и хорошо усваиваются. Искусственно же произведенные антиоксидантные препараты никогда не всасываются полностью. Многие продукты содержат антиоксиданты: фрукты, ягоды, красное вино, зеленый чай (подробнее см. в рамке на с. 150). Например, витамин С есть в растительных продуктах, но важно знать, что он разрушается при длительном хранении. Поэтому, когда заканчивается сезон свежих фруктов и овощей, нам стоит начать принимать витамин С в виде драже. Что касается пищевых добавок (БАД), научно проработанными являются только те препараты, которые продаются в аптеке. У каждого из них своя антиоксидантная активность, поэтому оптимальная доза и курс приема зависят от конкретной ситуации – с этим вопросом лучше обратиться к врачу.
Искусственно же произведенные антиоксидантные препараты никогда не всасываются полностью. Многие продукты содержат антиоксиданты: фрукты, ягоды, красное вино, зеленый чай (подробнее см. в рамке на с. 150). Например, витамин С есть в растительных продуктах, но важно знать, что он разрушается при длительном хранении. Поэтому, когда заканчивается сезон свежих фруктов и овощей, нам стоит начать принимать витамин С в виде драже. Что касается пищевых добавок (БАД), научно проработанными являются только те препараты, которые продаются в аптеке. У каждого из них своя антиоксидантная активность, поэтому оптимальная доза и курс приема зависят от конкретной ситуации – с этим вопросом лучше обратиться к врачу.
Могут ли антиоксиданты быть опасными?
С. Е.: По большей части это водорастворимые вещества, поэтому даже в большом количестве они безвредны и сами выводятся из организма. Однако не стоит долго употреблять (без контроля врача) пищевые добавки, механизм действия которых не изучен до конца. Они полезны лишь при повышенных нагрузках – в период эмоционального перенапряжения, при респираторных и вирусных заболеваниях, во время тяжелой сессии или сдачи сложного проекта на работе. Но в целом лучше рассчитывать на собственные силы организма, тем более что у нас есть все возможности укреплять их, тренировать наши приспособительные системы.
Они полезны лишь при повышенных нагрузках – в период эмоционального перенапряжения, при респираторных и вирусных заболеваниях, во время тяжелой сессии или сдачи сложного проекта на работе. Но в целом лучше рассчитывать на собственные силы организма, тем более что у нас есть все возможности укреплять их, тренировать наши приспособительные системы.
5 живых источников
| Акримекс | Р-р д/в/в и в/м введения 50 мг/1 мл: амп. 2 мл или 5 мл 5 или 10 шт. рег. №: ЛП-№(000245)-(РГ-R U) от 25. 05.21 Предыдущий рег. №: ЛП-006609 05.21 Предыдущий рег. №: ЛП-006609 | |||
| Акримекс | Таб., покр. пленочной оболочкой, 125 мг: 30, 36 или 50 шт. рег. №: ЛП-005475 от 18.04.19 Дата перерегистрации: 03.08.20 | |||
| Астрокс | Р-р д/в/в и в/м введения 50 мг/1 мл: амп. | |||
| Мексидол® | Р-р д/в/в и в/м введения 50 мг/1 мл: амп. 2 мл 5, 10, 20, 50 или 100 шт. или 5 мл 5, 10, 15, 20, 50 или 100 шт. рег. №: ЛП-№(000107)-(РГ-R U) от 29.12.20 Предыдущий рег. №: Р N002161/01 №: Р N002161/01 | |||
| Мексидол® | Таб., покр. пленочной оболочкой, 125 мг: 10, 20, 30, 40 или 50 шт. рег. №: ЛП-№(000086)-(РГ-R U) от 30.10.20 Предыдущий рег. №: ЛСР-002063/07 | |||
| Мексидол® Форте 250 | Таб. | |||
| Мексидониум | Р-р д/в/в и в/м введения 50 мг/1 мл: амп. 2 мл или 5 мл 5 или 10 шт. рег. №: ЛП-006793 от 18.02.21 | |||
| Мексикор® | Капс. | |||
| Мексикор® | Р-р д/в/в и в/м введения 50 мг/1 мл: 2 мл амп. 10 шт.; 5 мл амп. 5, 10 или 20 шт. рег. №: Р N002948/01 от 22. 09.08
Дата перерегистрации: 24.04.14 09.08
Дата перерегистрации: 24.04.14 | |||
| Мексилек-Лекфарм | Р-р д/в/в и в/м введения 50 мг/мл: 2 мл амп. 5 или 10 шт. рег. №: ЛП-004800 от 13.04.18 | |||
| Мексиприм® | Таб. | |||
| Мексипровел | Р-р д/в/в и в/м введения 50 мг/мл: 2 мл или 5 мл амп. 5 или 10 шт. рег. №: ЛП-006794 от 18.02.21 | |||
| Мекситерра | Таб. | |||
| Мексифин | Р-р д/в/в и в/м введения 100 мг/2 мл: амп. 10 или 20 шт. рег. №: ЛСР-001967/07 от 07.08.07 | |||
| Мексифин | Р-р д/в/в и в/м введения 250 мг/5 мл: амп. | |||
| МЕКСэллара® | Р-р д/в/в и в/м введения 50 мг/мл: 2 мл или 5 мл амп. 5 или 10 шт. рег. №: ЛП-005620 от 01.07.19 | |||
| Метостабил | Р-р д/в/в и в/м введения 50 мг/мл: амп. | Произведено: ОЗОН (Россия) | ||
| Метостабил | Таб., покр. оболочкой, 125 мг: 5, 10, 15, 20, 25, 30, 40, 50, 60, 75, 80, 90, 100, 120, 125, 150, 180, 200, 240, 250 или 300 шт. | Произведено: ОЗОН (Россия) | ||
| Метуцинвел | Р-р д/в/в и в/м введения 50 мг/мл: 2 мл или 5 мл амп. 5 или 10 шт. | |||
| Наномексил | Р-р д/в/в и в/м введения 50 мг/мл: 2 мл или 5 мл амп. 5 или 10 шт. рег. №: ЛП-006797 от 18.02.21 | |||
| Нейрокард | Р-р д/в/в и в/м введения 50 мг/мл: 2 мл или 5 мл амп. | |||
| Нейрокс® | Р-р д/в/в и в/м введения 100 мг/2 мл: амп. 5, 10, 20 или 50 шт. рег. №: ЛП-№(000316)-(РГ-R U) от 20.07.21 Предыдущий рег. №: ЛСР-007439/09 №: ЛСР-007439/09Р-р д/в/в и в/м введения 250 мг/5 мл: амп. 5, 10 или 20 шт. рег. №: ЛП-№(000316)-(РГ-R U) от 20.07.21 Предыдущий рег. №: ЛСР-007439/09 | |||
| Нейрокс® | Таб., покр. пленочной оболочкой, 125 мг: 10, 30 или 50 шт. рег. №: ЛП-005670 от 22. 07.19 07.19 | |||
| Нейромексол® | Таб., покр. пленочной оболочкой, 125 мг: 30 или 50 шт. рег. №: ЛП-006028 от 13.01.20 Дата перерегистрации: 16.09.21 | |||
| Проинин® | Р-р д/в/в и в/м введения 50 мг/мл: 2мл или 5 мл амп. | |||
| Церекард | Р-р д/в/в и в/м введения 100 мг/2 мл: амп. 5 или 10 шт. рег. №: ЛСР-003013/09 от 16.04.09 | |||
| Церекард | Р-р д/в/в и в/м введения 250 мг/5 мл: амп. | |||
| Цитореан® | Таб., покр. пленочной оболочкой, 125 мг: 60, 90 или 120 шт. рег. №: ЛП-006076 от 05.02.20 | |||
| Эврин | Р-р д/в/в и в/м введения 50 мг/1 мл: амп. | |||
| Этилметилгидроксипиридина сукцинат | Р-р д/в/в и в/м введения 50 мг/мл: 2 мл амп. 10 шт или 5 мл амп. 5 шт. рег. №: ЛП-006560 от 11.11.20 | Произведено: ОЗОН (Россия) | ||
| Этилметилгидроксипиридина сукцинат | Р-р д/в/в и в/м введения 50 мг/мл: 2 мл, 5 мл или 10 мл амп. | |||
 2 мл 10, 100, 250 или 500 шт.
2 мл 10, 100, 250 или 500 шт. , покр. пленочной оболочкой, 250 мг: 10, 20, 30, 40, 50 или 60 шт.
, покр. пленочной оболочкой, 250 мг: 10, 20, 30, 40, 50 или 60 шт. 100 мг: 20, 30 или 60 шт.
100 мг: 20, 30 или 60 шт. , покр. пленочной оболочкой, 125 мг: 10, 20, 30, 40 или 60 шт.
, покр. пленочной оболочкой, 125 мг: 10, 20, 30, 40 или 60 шт. , покр. пленочной оболочкой, 125 мг: 10, 20, 30, 40 или 50 шт.
, покр. пленочной оболочкой, 125 мг: 10, 20, 30, 40 или 50 шт.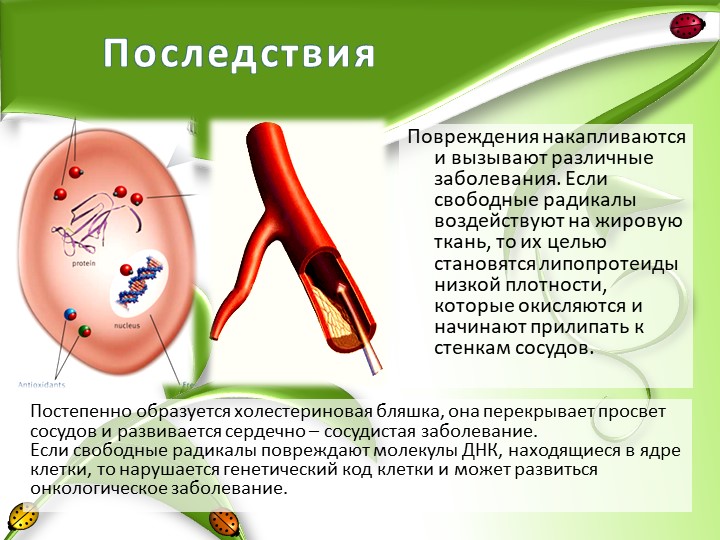 10 или 20 шт.
10 или 20 шт. 5 или 10 шт.
5 или 10 шт.
 5 или 10 шт.
5 или 10 шт. 5 или 10 шт.
5 или 10 шт. 5 или 10 шт.
5 или 10 шт. 2 мл 10 или 20 шт., 5 мл 5 или 20 шт.
2 мл 10 или 20 шт., 5 мл 5 или 20 шт.Понимание оксидантов и антиоксидантов: классическая команда с новыми игроками
Свободнорадикальные окислители, такие как активные формы кислорода, реактивные формы азота и реактивные формы серы, вырабатываются внутри клеток посредством различных метаболических процессов. Организм оснащен системой антиоксидантной защиты, которая защищает от окислительного повреждения, вызванного этими реактивными окислителями, и играет важную роль в защите клеток от окислительного стресса и повреждений.Антиоксиданты, такие как глутатион (GSH), тиоредоксин, аскорбиновая кислота и ферменты, например, супероксиддисмутаза (SOD), глутатионпероксидаза (GPx), каталаза (CAT), противодействуют окислительному стрессу и защищают липиды, белки и ДНК. Антиоксиданты, такие как токоферолы, аскорбиновая кислота, каротиноиды, флавоноиды, аминокислоты, также являются природными антиоксидантами, присутствующими в пищевых продуктах. Растет спрос и доступность дизайнерских продуктов, обогащенных антиоксидантами и пробиотиками, которые могут быть важны для здоровья человека.В обзорной статье представлен краткий обзор оксидантов и антиоксидантных систем в организме человека, включая роль пробиотиков и воспаления. ПРАКТИЧЕСКОЕ ПРИМЕНЕНИЕ: Антиоксиданты, такие как GSH, тиоредоксин, аскорбиновая кислота и др., и защитные ферменты, например, SOD, GPx, CAT и др. противодействуют окислительному стрессу и защищают клеточные биомолекулы. Антиоксиданты, такие как токоферолы, аскорбиновая кислота, каротиноиды, флавоноиды, аминокислоты, фосфолипиды и стеролы, являются естественными антиоксидантами, содержащимися в потребляемых продуктах.Они играют важную роль в удалении свободных радикалов и нерадикальных оксидантов и защищают клетки от окислительного стресса и повреждений. Важность антиоксидантов можно понять из того факта, что окислительное повреждение в настоящее время связано с целым рядом заболеваний, включая рак, нейродегенерацию, диабет и т. д. В некоторых подходах к улучшению здоровья человека и достижению долголетия пищевые антиоксиданты используются в составе диетических и обогащенных пищевых продуктов. Антиоксиданты также сохраняют свежесть и продлевают срок годности пищевых продуктов.Обогащенные или дизайнерские продукты, в которые добавлены антиоксидантные питательные вещества и использование микроорганизмов в качестве пробиотиков, становятся все более доступными на рынке в качестве здоровой пищи и пищевых добавок.
Важность антиоксидантов можно понять из того факта, что окислительное повреждение в настоящее время связано с целым рядом заболеваний, включая рак, нейродегенерацию, диабет и т. д. В некоторых подходах к улучшению здоровья человека и достижению долголетия пищевые антиоксиданты используются в составе диетических и обогащенных пищевых продуктов. Антиоксиданты также сохраняют свежесть и продлевают срок годности пищевых продуктов.Обогащенные или дизайнерские продукты, в которые добавлены антиоксидантные питательные вещества и использование микроорганизмов в качестве пробиотиков, становятся все более доступными на рынке в качестве здоровой пищи и пищевых добавок.
Ключевые слова: антиоксиданты; ферментативные антиоксиданты; свободные радикалы; функциональные продукты; натуральные антиоксиданты; окислители; пробиотики.
Оксиданты и антиоксиданты: друзья или враги?
Хотя существование «свободных радикалов» было известно в области химии уже давно, эти интересные виды окисляющих молекул привлекли внимание ученых-медиков и врачей в начале 1950-х годов, когда Денхэм Харман начал публиковать ряд сообщает о «свободнорадикальной теории старения» [1]. Два десятилетия спустя, основанная на признании того, что образование свободных радикалов в клетке происходит в основном в митохондриях и что мутации митохондриальной ДНК (мтДНК) активно участвуют в процессе старения, эта теория превратилась в «митохондриальную теорию старения». 2, 3]. Несмотря на ярлык «теория», весь медицинский мир верит, по крайней мере частично, в истинность этого объяснения основных механизмов — неизбежного биологического процесса — старения.
Два десятилетия спустя, основанная на признании того, что образование свободных радикалов в клетке происходит в основном в митохондриях и что мутации митохондриальной ДНК (мтДНК) активно участвуют в процессе старения, эта теория превратилась в «митохондриальную теорию старения». 2, 3]. Несмотря на ярлык «теория», весь медицинский мир верит, по крайней мере частично, в истинность этого объяснения основных механизмов — неизбежного биологического процесса — старения.
Начиная с 1970-х годов в медицинской литературе стали появляться обширные исследования, выясняющие связь свободных радикалов с тем или иным патофизиологическим состоянием, что привело к формулировке определения «свободнорадикальные заболевания» [4].Благодаря этому многие патологии, такие как гипертоническая болезнь, атеросклероз, аутоиммунные заболевания и рак, были объяснены участием свободных радикалов [5]. В зависимости от молекулярного источника или основы простой термин «свободный радикал» расширился за счет других описаний, таких как «активные формы кислорода» или «свободные радикалы кислорода». Вскоре после открытия Ignarro et al. [6] эндогенным сосудорасширяющим медиатором, широко известным как релаксирующий фактор эндотелия (EDRF), был оксид азота (NO • ), газообразная радикальная молекула, другой термин, а именно «реактивный формы азота» были включены в номенклатуру этой конкретной области науки.
Вскоре после открытия Ignarro et al. [6] эндогенным сосудорасширяющим медиатором, широко известным как релаксирующий фактор эндотелия (EDRF), был оксид азота (NO • ), газообразная радикальная молекула, другой термин, а именно «реактивный формы азота» были включены в номенклатуру этой конкретной области науки.
Значительное количество радикалов, таких как супероксидный свободный радикал-анион (O 2 •− ) или гидроксильный радикал ( • OH), а также другая группа «нерадикальных реакционноспособных молекул», таких как перекись водорода ( H 2 O 2 ) и пероксинитрит (ONOO − ), и в последние десятилетия были описаны различные вредные эффекты этих молекул [7, 8]. В конечном итоге повреждение клеток этими высокореакционноспособными формами кислорода и азота (АФК и АФК) происходит в результате изменения макромолекул [9, 10].К ним относятся липопероксидация полиненасыщенных жирных кислот в мембранных липидах, окисление белков, разрыв цепи ДНК [11–14], окисление РНК [15], митохондриальная деполяризация и апоптоз. Мутации ядерного белка р53, которые могут привести к апоптозу, также связаны с окислительным стрессом. Нарушения клеточных/тканевых функций, вызванные окислительным стрессом, связаны с болезненными состояниями, а именно. , болезнь Альцгеймера [16] и болезнь Паркинсона [17], различные виды рака [18] и процессы старения [19] среди прочих.В нормальных условиях химически активные частицы удаляются антиоксидантами, которые, в широком смысле, относятся к молекулам, способным напрямую вступать в реакцию с окислителями для снижения их окислительной способности, например. очищающие ферменты, такие как супероксиддисмутаза, каталаза, глутатионпероксидаза, и т.д. полифенолов. Эти молекулы могут быть природными или синтетическими, либо гидрофильными, такими как аскорбиновая кислота, либо гидрофобными, такими как α-токоферол.Благодаря этим действиям антиоксиданты могут либо предотвращать образование окисляющих соединений, либо уменьшать действие опасных метаболических или ксенобиотических оксидантов и, следовательно, предотвращать возникновение острых или хронических заболеваний в организме и/или восстанавливать уже нанесенные повреждения клеток/тканей.
Мутации ядерного белка р53, которые могут привести к апоптозу, также связаны с окислительным стрессом. Нарушения клеточных/тканевых функций, вызванные окислительным стрессом, связаны с болезненными состояниями, а именно. , болезнь Альцгеймера [16] и болезнь Паркинсона [17], различные виды рака [18] и процессы старения [19] среди прочих.В нормальных условиях химически активные частицы удаляются антиоксидантами, которые, в широком смысле, относятся к молекулам, способным напрямую вступать в реакцию с окислителями для снижения их окислительной способности, например. очищающие ферменты, такие как супероксиддисмутаза, каталаза, глутатионпероксидаза, и т.д. полифенолов. Эти молекулы могут быть природными или синтетическими, либо гидрофильными, такими как аскорбиновая кислота, либо гидрофобными, такими как α-токоферол.Благодаря этим действиям антиоксиданты могут либо предотвращать образование окисляющих соединений, либо уменьшать действие опасных метаболических или ксенобиотических оксидантов и, следовательно, предотвращать возникновение острых или хронических заболеваний в организме и/или восстанавливать уже нанесенные повреждения клеток/тканей. Поэтому неудивительно, что большое количество исследований было сосредоточено на молекулах с антиоксидантной активностью в терапевтических целях для противодействия вредному воздействию реактивных частиц и окислительного стресса. Однако следует отметить, что вместо этого было доказано, что значительное количество молекул антиоксидантов обладает прооксидантным потенциалом и способствует окислительным реакциям [20].Таким образом, использование антиоксидантов для профилактики возможных радикальных поражений, а именно «антиоксидантная терапия», на сегодняшний день все еще остается спорным вопросом [21, 22] и может объяснить часто противоречивые результаты в исследованиях на людях.
Поэтому неудивительно, что большое количество исследований было сосредоточено на молекулах с антиоксидантной активностью в терапевтических целях для противодействия вредному воздействию реактивных частиц и окислительного стресса. Однако следует отметить, что вместо этого было доказано, что значительное количество молекул антиоксидантов обладает прооксидантным потенциалом и способствует окислительным реакциям [20].Таким образом, использование антиоксидантов для профилактики возможных радикальных поражений, а именно «антиоксидантная терапия», на сегодняшний день все еще остается спорным вопросом [21, 22] и может объяснить часто противоречивые результаты в исследованиях на людях.
С другой стороны, также стало очевидным, что свободные радикалы не только участвуют в патологических процессах, но и их существование необходимо для многих физиологических функций живых организмов [23, 24], в том числе для «здорового старения» [25, 26 ]. Сообщалось также, что перекисное окисление липидов, основное последствие повреждения свободными радикалами, может иметь как вредные, так и положительные эффекты [27, 28]. В настоящее время широко известно, что эти биологически «гиперактивные» молекулы действуют как сигнальные агенты в различных клеточных путях, открывая новую эру исследований, так называемую «окислительно-восстановительную передачу сигналов» [29–31]. Перекись водорода и пероксинитрит, в частности, вовлечены в значительное число клеточных сигнальных каскадов [32–34]; в зависимости от их нерадикальной структуры эти молекулы имеют относительно более длительный период полураспада, чем почти все другие окислители, что позволяет им мигрировать от мест их образования и диффундировать через мембраны.При этом сообщалось, что факторы транскрипции, такие как AP-1, NF-κB и/или Nrf2, участвуют в этих редокс-модулируемых сигнальных путях [35–37].
В настоящее время широко известно, что эти биологически «гиперактивные» молекулы действуют как сигнальные агенты в различных клеточных путях, открывая новую эру исследований, так называемую «окислительно-восстановительную передачу сигналов» [29–31]. Перекись водорода и пероксинитрит, в частности, вовлечены в значительное число клеточных сигнальных каскадов [32–34]; в зависимости от их нерадикальной структуры эти молекулы имеют относительно более длительный период полураспада, чем почти все другие окислители, что позволяет им мигрировать от мест их образования и диффундировать через мембраны.При этом сообщалось, что факторы транскрипции, такие как AP-1, NF-κB и/или Nrf2, участвуют в этих редокс-модулируемых сигнальных путях [35–37].
В совокупности текущий консенсус заключается в том, что контролируемое и устойчивое производство как радикальных, так и нерадикальных реакционноспособных молекул необходимо для нормальных физиологических и клеточных функций; однако их неконтролируемое или чрезмерное производство может вызвать «окислительный/нитрозативный стресс», приводящий к разрушению структурных биомолекул, что приводит к клеточной дисфункции и гибели и, в конечном итоге, к повреждению или отказу тканей и органов. Научный мир активно занимается исследованием того, являются ли оксиданты или антиоксиданты друзьями или врагами друг для друга и/или для живых организмов; проводится все больше исследований с целью выяснения механизмов действия эндогенно продуцируемых окисляющих молекул, их связи с физиологическими процессами и взаимодействия с другими биомолекулами.
Научный мир активно занимается исследованием того, являются ли оксиданты или антиоксиданты друзьями или врагами друг для друга и/или для живых организмов; проводится все больше исследований с целью выяснения механизмов действия эндогенно продуцируемых окисляющих молекул, их связи с физиологическими процессами и взаимодействия с другими биомолекулами.
В этом первом выпуске « Оксиданты и антиоксиданты в медицинской науке » мы объявляем о новом периодическом ресурсе для специалистов-исследователей в этой привлекательной области, чтобы найти возможность поделиться своим опытом и знаниями с медицинскими работниками через медицинский литература.Мы начинаем с выпуска из 10 статей от авторов и исследовательских групп со всего мира, включая США, Кубу, Бельгию, Эстонию, Индию и Малайзию. В первый выпуск вошли как обзорные статьи, авторизованные старшими академиками, так и научные статьи в области окислительно-восстановительной науки. Наша главная цель — предоставить актуальные и надежные знания ученым в этой области и, таким образом, открыть новую дверь в мир и секреты оксидантов и антиоксидантов в медицинской науке.
Оксиданты, антиоксиданты и благотворное влияние образования реактивных частиц, вызванного физическими упражнениями
В этом обзоре представлен обзор влияния реактивных соединений, образующихся во время упражнений, и их влияние на адаптацию к физической нагрузке.Реактивные частицы и свободные радикалы представляют собой нестабильные молекулы, которые окисляют другие молекулы, чтобы стать стабильными. Хотя они играют важную роль в нашем организме, они также могут привести к окислительному стрессу, нарушающему различные клеточные функции. Во время физической нагрузки реактивные частицы могут образовываться в основном, но не исключительно, за счет следующих механизмов: утечка электронов в митохондриальной цепи переноса электронов, ишемия/реперфузия и активация эндотелиальной ксантиноксидазы, воспалительная реакция и аутоокисление катехоламинов.Постоянные упражнения также приводят к усилению антиоксидантной защиты организма, что помогает свести к минимуму окислительный стресс, который может возникнуть после интенсивной тренировки. Недавние исследования показывают полезную роль реактивных частиц, образующихся во время тренировки, которые приводят к важным тренировочным адаптациям: ангиогенезу, биогенезу митохондрий и мышечной гипертрофии. Адаптация происходит в зависимости от механического и, следовательно, биохимического стимула внутри мышцы. Это новая область исследований, которая обещает важные открытия в области молекулярных и клеточных механизмов, связанных с взаимосвязью между окислительным стрессом и физическими упражнениями.
Недавние исследования показывают полезную роль реактивных частиц, образующихся во время тренировки, которые приводят к важным тренировочным адаптациям: ангиогенезу, биогенезу митохондрий и мышечной гипертрофии. Адаптация происходит в зависимости от механического и, следовательно, биохимического стимула внутри мышцы. Это новая область исследований, которая обещает важные открытия в области молекулярных и клеточных механизмов, связанных с взаимосвязью между окислительным стрессом и физическими упражнениями.
1. Введение
В последние годы был достигнут значительный прогресс в окислительно-восстановительной биологии упражнений, что делает этот обзор весьма актуальным для области спортивной науки. Исследования показали, что физические упражнения увеличивают производство реактивных частиц и свободных радикалов. Хотя в избытке дисбаланс между этими молекулами оксидантов и антиоксидантами в организме может привести к пагубным последствиям для нашего организма, как показывают исследования, реактивные частицы, образующиеся во время упражнений, играют важную роль в адаптации мышц к упражнениям.
Этот обзор начинается с объяснения различных режимов упражнений, а затем предлагает обзор производства реактивных частиц, вызванных физическими упражнениями. Далее следует описание антиоксидантной сети организма, а также конкретные сведения о ферментативных и неферментативных антиоксидантах. Обсуждается также эффективность пищевых антиоксидантных добавок. Наконец, в обзоре представлены важные данные о том, как реактивные виды во время упражнений приводят к благоприятной адаптации, вызванной физическими упражнениями, такой как усиление активности антиоксидантных ферментов, ангиогенез, биогенез митохондрий и мышечная гипертрофия.Наш подход обеспечивает общее понимание предмета и представляет новые интересные результаты; при необходимости делается ссылка на более конкретные обзоры.
2. Определение упражнения
Упражнение можно определить как любую запланированную структурированную деятельность, которая приводит к увеличению расхода энергии и частоты сердечных сокращений.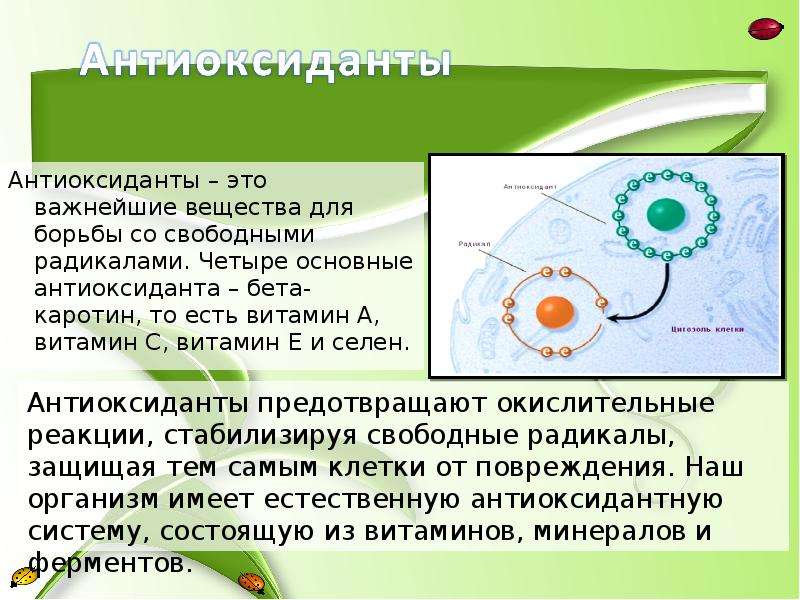 Существуют различные режимы упражнений в зависимости от интенсивности (аэробные и анаэробные), сокращения мышц (изометрические, концентрические и эксцентрические) и частоты (острые и хронические).Необходимо объяснить каждый из этих режимов упражнений, чтобы читатель мог лучше понять влияние упражнений на производство реактивных видов и его последствия для ангиогенеза, гипертрофии и биогенеза митохондрий.
Существуют различные режимы упражнений в зависимости от интенсивности (аэробные и анаэробные), сокращения мышц (изометрические, концентрические и эксцентрические) и частоты (острые и хронические).Необходимо объяснить каждый из этих режимов упражнений, чтобы читатель мог лучше понять влияние упражнений на производство реактивных видов и его последствия для ангиогенеза, гипертрофии и биогенеза митохондрий.
2.1. Различия между аэробными и анаэробными упражнениями
Основное физиологическое различие между аэробными и анаэробными упражнениями заключается в источнике энергии. С одной стороны, аэробные упражнения можно охарактеризовать использованием аэробного метаболизма во время физической нагрузки.В этом случае аэробный метаболизм преимущественно вырабатывает энергию из жира, а с использованием кислорода вырабатывает энергию без большого накопления молочной кислоты в крови. С другой стороны, анаэробные упражнения характеризуются короткими периодами усилий от высоких до максимальных, когда энергия поступает за счет анаэробного метаболизма, то есть без использования кислорода, что приводит к высокому накоплению молочной кислоты в крови [1, 2]. Примерами упражнений на аэробную выносливость являются бег трусцой, бег, плавание, гребля и езда на велосипеде, когда они выполняются примерно на 50–75% от максимальной аэробной способности участников (VO2max) и в течение относительно продолжительных периодов времени.Примерами анаэробных упражнений являются спринты (во время плавания, езды на велосипеде или бега), прыжки в длину, соревновательная тяжелая атлетика или любые другие упражнения продолжительностью до 2 минут, выполняемые с превышением лактатного порога участников (например, >75% VO2max для тренированных людей).
Примерами упражнений на аэробную выносливость являются бег трусцой, бег, плавание, гребля и езда на велосипеде, когда они выполняются примерно на 50–75% от максимальной аэробной способности участников (VO2max) и в течение относительно продолжительных периодов времени.Примерами анаэробных упражнений являются спринты (во время плавания, езды на велосипеде или бега), прыжки в длину, соревновательная тяжелая атлетика или любые другие упражнения продолжительностью до 2 минут, выполняемые с превышением лактатного порога участников (например, >75% VO2max для тренированных людей).
2.2. Различия между изометрическими, концентрическими и эксцентрическими упражнениями
Сокращение мышц происходит, когда центральная нервная система передает сигнал мышечным волокнам. При наличии достаточного количества энергии и кальция мышечные волокна создают напряжение, и мышца может укорачиваться, удлиняться или оставаться прежнего размера.Концентрическое сокращение относится к укорочению мышцы с созданием силы. Примеры концентрических упражнений включают фазу скручивания приседаний, когда два конца (начало и место прикрепления) мышц живота сближаются, сокращаются и укорачиваются [1, 3]. Эксцентрическое сокращение соответствует «растягиванию» мышцы из концентрического или статического положения при напряжении. Примером эксцентрического упражнения является фаза опускания сгибания рук на бицепс, когда противоположная сила, создаваемая гантелью, создает силу, большую или равную силе, создаваемой для удлинения двух концов бицепса в исходное положение [1].Изометрическое упражнение состоит в сокращении мышц, происходящем без движения в пораженных суставах, например, когда спортсмен держит штангу в статике под углом 90 градусов в упражнении на бицепс.
Примеры концентрических упражнений включают фазу скручивания приседаний, когда два конца (начало и место прикрепления) мышц живота сближаются, сокращаются и укорачиваются [1, 3]. Эксцентрическое сокращение соответствует «растягиванию» мышцы из концентрического или статического положения при напряжении. Примером эксцентрического упражнения является фаза опускания сгибания рук на бицепс, когда противоположная сила, создаваемая гантелью, создает силу, большую или равную силе, создаваемой для удлинения двух концов бицепса в исходное положение [1].Изометрическое упражнение состоит в сокращении мышц, происходящем без движения в пораженных суставах, например, когда спортсмен держит штангу в статике под углом 90 градусов в упражнении на бицепс.
2.3. Различия между хронической и острой физической нагрузкой
Острая физическая нагрузка может характеризоваться единичными приступами физической нагрузки, вызывающими временные метаболические и сердечно-сосудистые реакции продолжительностью от нескольких минут до нескольких часов. Хронические упражнения — это повторные серии упражнений, вызывающие тренировочную реакцию, когда физиологические и метаболические адаптации становятся более заметными и продолжительными [4].Как аэробные, так и анаэробные упражнения могут выполняться либо постоянно, либо остро.
Хронические упражнения — это повторные серии упражнений, вызывающие тренировочную реакцию, когда физиологические и метаболические адаптации становятся более заметными и продолжительными [4].Как аэробные, так и анаэробные упражнения могут выполняться либо постоянно, либо остро.
2.4. Компоненты физической подготовки
Компоненты физической подготовки включают в себя продолжительность, частоту, интенсивность и тип деятельности программы тренировок; эти переменные необходимо тщательно рассмотреть, чтобы удостовериться, что цели программы успешно достигнуты. Важно отметить, что тренировочные стратегии для улучшения, например, аэробной производительности отличаются от стратегий, необходимых для улучшения силовой подготовленности, и у каждого вида деятельности есть особые потребности, которые будут влиять на использование определенных компонентов физической подготовки [1].Тип спорта или активности будет определять, какие упражнения следует включать, чтобы обеспечить идеальное улучшение физиологических показателей. Аэробные тренировки настоятельно рекомендуются для развития беговых качеств на длинные дистанции, но тот же план тренировок не применим для соревнований по тяжелой атлетике. Поэтому важно определиться с видом спорта, прежде чем применять компоненты фитнеса в тренировках.
Аэробные тренировки настоятельно рекомендуются для развития беговых качеств на длинные дистанции, но тот же план тренировок не применим для соревнований по тяжелой атлетике. Поэтому важно определиться с видом спорта, прежде чем применять компоненты фитнеса в тренировках.
Также очень важно тренироваться с такой частотой, которая обеспечит стимул, вызывающий положительную физиологическую адаптацию, и уравновешивать его достаточным временем восстановления для повышения производительности.Некоторые виды деятельности потребуют большей частоты, чем другие, что компенсируется корректировкой интенсивности упражнений. Интенсивность определяется тренировочной целью и текущим уровнем физической подготовки спортсмена. Этим манипулируют, например, работая с разным процентом от их максимальной аэробной способности (%VO2max) или максимума повторений (1RM), обычно прогрессивным образом, в то время как время (продолжительность) тренировочной сессии должно контролироваться в зависимости от интенсивности [1, 3]. Увеличение продолжительности сеанса упражнений влияет на компонент физической подготовки и, при правильном управлении, может привести к положительному увеличению, среди прочего, производительности.
Увеличение продолжительности сеанса упражнений влияет на компонент физической подготовки и, при правильном управлении, может привести к положительному увеличению, среди прочего, производительности.
В разных видах спорта и занятиях используются разные энергетические системы и задействуются разные мышечные волокна, поэтому требуются разные тренировочные стратегии, манипулирующие компонентами фитнеса. Бегунам на короткие дистанции потребуется более высокая анаэробная способность и интенсивность упражнений, а также меньшее количество упражнений по сравнению с бегунами на длинные дистанции. Кроме того, такие виды спорта, как теннис, включают эксцентрические движения нижней части тела, поэтому предписание упражнений для этого должно быть направлено на улучшение эксцентрической силы за счет использования таких же движений и интенсивности, как и в игре.Таким образом, становится ясно, что компоненты физической подготовки взаимосвязаны, и комбинированное воздействие на них всех необходимо для улучшения физиологических показателей [1].
В этом обзоре будут подробно обсуждаться эффекты аэробных и анаэробных упражнений при их интенсивном или постоянном выполнении, а также различия между выработкой реактивных частиц в упражнениях, использующих различное сокращение мышц.
3. Производство реактивных частиц во время учений
3.1. Реакционноспособные соединения и свободные радикалы
Реактивные соединения и свободные радикалы представляют собой молекулы, которые из-за своей молекулярной нестабильности (например, неспаренный электрон) способствуют реакциям окисления с другими молекулами, такими как белки, липиды и ДНК, чтобы стать стабилизированный [5–7]. Многие реакционноспособные соединения являются кислородоцентрированными (O 3 , H 2 O 2 и т. д.) и, таким образом, называются активными формами кислорода (АФК). Некоторые АФК также являются свободными радикалами, такие как анион супероксида (O2-) и оксид азота (NO), поскольку они имеют неспаренный электрон [6]. Свободные радикалы и реактивные формы необходимы для нашего благополучия, играя различные регулирующие роли в клетках. Например, АФК продуцируются иммунными клетками — нейтрофилами и макрофагами — в процессе респираторного взрыва с целью элиминации антигенов [8]. Они также служат стимулирующими сигналами нескольких генов, которые кодируют факторы транскрипции, дифференцировки и развития, а также стимулируют межклеточную адгезию, передачу клеточных сигналов, участие в вазорегуляции и пролиферации фибробластов, а также повышенную экспрессию антиоксидантных ферментов [9, 10].Последний пример наблюдается, когда люди постоянно выполняют физические упражнения, и будет обсуждаться далее в разделе 4.
Свободные радикалы и реактивные формы необходимы для нашего благополучия, играя различные регулирующие роли в клетках. Например, АФК продуцируются иммунными клетками — нейтрофилами и макрофагами — в процессе респираторного взрыва с целью элиминации антигенов [8]. Они также служат стимулирующими сигналами нескольких генов, которые кодируют факторы транскрипции, дифференцировки и развития, а также стимулируют межклеточную адгезию, передачу клеточных сигналов, участие в вазорегуляции и пролиферации фибробластов, а также повышенную экспрессию антиоксидантных ферментов [9, 10].Последний пример наблюдается, когда люди постоянно выполняют физические упражнения, и будет обсуждаться далее в разделе 4.
Наше тело имеет сложную сеть антиоксидантов, которая действует как защитная система, нейтрализующая свободные радикалы и химически активные частицы. Этот процесс позволяет поддерживать гомеостаз. Тем не менее, окислительный стресс и нарушение клеточной функции могут возникать, если происходит резкое увеличение концентрации оксидантов в организме, подавляющее доступные антиоксиданты. Это также может произойти при истощении доступных антиоксидантов из-за болезни или неправильного питания [11]. На самом деле хронический окислительный стресс считается причиной или следствием многих острых и хронических заболеваний человека [7, 12, 13], например, ожирения, сердечно-сосудистых заболеваний, рака, острого повреждения легких и рассеянного склероза.
Это также может произойти при истощении доступных антиоксидантов из-за болезни или неправильного питания [11]. На самом деле хронический окислительный стресс считается причиной или следствием многих острых и хронических заболеваний человека [7, 12, 13], например, ожирения, сердечно-сосудистых заболеваний, рака, острого повреждения легких и рассеянного склероза.
Непосредственно измерить образование свободных радикалов и реактивных частиц очень сложно из-за их высокой реакционной способности и низкой стационарной концентрации.Тем не менее, метод электронного спинового резонанса (ЭПР) является прямым методом обнаружения частиц, которые имеют неспаренный электрон, что обычно означает, что это свободный радикал [14]. Однако этот метод и несколько других прямых измерений требуют очень дорогого оборудования и имеют сложные методы. Таким образом, для оценки оксидативного стресса в основном используются косвенные методы. Для измерения биомаркеров окислительного стресса в мышечной ткани человека можно использовать выдыхаемый воздух, мочу, кровь и промывание носа [15–19]. Согласно Пауэрсу и Джексону [10], надежные маркеры этого процесса должны обладать следующими характеристиками: быть химически уникальными, химически обнаруживаемыми и иметь относительно длительный период полураспада, увеличиваться или уменьшаться в процессе окислительного стресса и не подвергаться влиянию другие клеточные циклы, чтобы избежать смешанных факторов. Кроме того, окислительный стресс также обычно измеряют, анализируя сдвиг в антиоксидантной системе организма и активность специфических антиоксидантных ферментов [5].
Согласно Пауэрсу и Джексону [10], надежные маркеры этого процесса должны обладать следующими характеристиками: быть химически уникальными, химически обнаруживаемыми и иметь относительно длительный период полураспада, увеличиваться или уменьшаться в процессе окислительного стресса и не подвергаться влиянию другие клеточные циклы, чтобы избежать смешанных факторов. Кроме того, окислительный стресс также обычно измеряют, анализируя сдвиг в антиоксидантной системе организма и активность специфических антиоксидантных ферментов [5].
3.2. Окислительный стресс, вызванный физической нагрузкой
В нормальных условиях скелетная мышца вырабатывает анионы супероксида и NO с низкой скоростью. Однако во время сократительной деятельности эта скорость резко увеличивается. На самом деле аэробные упражнения связаны с увеличением потребления кислорода как всем телом, так и особенно сокращающейся группой мышц. Sen [20] сообщил об увеличении в 10–15 раз скорости потребления кислорода всем телом и увеличении более чем в 100 раз потока кислорода в активных мышцах во время аэробных упражнений для всего тела.
Хотя прямых доказательств продукции АФК во время упражнений мало [14, 21, 22], главным образом из-за ограниченности используемых методов, существует множество литературы, косвенно подтверждающей, что окислительный стресс может возникать во время аэробных и анаэробных упражнений. (обзор окислительного стресса, вызванного физическими упражнениями, см. у Fisher-Wellman и Bloomer [5]). Анализируя доступную литературу, можно сделать несколько разных выводов относительно уровня окислительного стресса в результате физической нагрузки.Это понятно, потому что на скорость окисления могут влиять различные факторы, такие как задействованные группы мышц, способы сокращения, интенсивность упражнений, продолжительность упражнений и тренирующаяся популяция. Поэтому используемые в исследованиях модели упражнений заслуживают критической оценки.
3.3. Механизмы увеличения образования свободных радикалов при физических нагрузках
Причины увеличения образования свободных радикалов и АФК во время упражнений до конца не выяснены. Хотя были идентифицированы различные механизмы, до сих пор отсутствует понимание того, как каждый из них способствует общему количеству вызванного окислительного стресса. Кроме того, эти механизмы могут действовать синергетически, и разные виды упражнений, вероятно, вызывают разные пути образования свободных радикалов [23]. Например, хотя общее мнение состоит в том, что во время упражнений выработка реактивных частиц происходит в основном за счет сокращения мышц (скелетных и сердечных), другие механизмы, такие как воспалительные процессы и повышенное высвобождение катехоламинов, которые могут возникать при физических нагрузках, также играют важную роль. важную роль в образовании реактивных частиц.Основные механизмы образования реактивных частиц описаны ниже.
Хотя были идентифицированы различные механизмы, до сих пор отсутствует понимание того, как каждый из них способствует общему количеству вызванного окислительного стресса. Кроме того, эти механизмы могут действовать синергетически, и разные виды упражнений, вероятно, вызывают разные пути образования свободных радикалов [23]. Например, хотя общее мнение состоит в том, что во время упражнений выработка реактивных частиц происходит в основном за счет сокращения мышц (скелетных и сердечных), другие механизмы, такие как воспалительные процессы и повышенное высвобождение катехоламинов, которые могут возникать при физических нагрузках, также играют важную роль. важную роль в образовании реактивных частиц.Основные механизмы образования реактивных частиц описаны ниже.
Утечка электронов в митохондриальной электрон-транспортной цепи
Эта теория привела к интерпретации того, что во время физических упражнений следует ожидать значительного увеличения образования свободных радикалов из-за «утечки» электронов в дыхательной цепи через внутреннюю мембрану митохондрий сокращающихся мышечных клеток. Это может происходить из-за неадекватной связи переноса электрона между комплексами I и III (рис. 1) [23, 24].По-видимому, комплекс I, а именно железо-серные кластеры, высвобождает активные формы кислорода — анион супероксида — только в направлении митохондриального матрикса, тогда как комплекс III — сайт окисления убихинола — высвобождает супероксид как в матрикс, так и за пределы внутренней мембраны [24]. , 25]. Несмотря на эту теоретическую привлекательность, существует мало прямых доказательств того, что производство митохондриальных супероксидных анионов увеличивается во время физических упражнений. Кроме того, есть исследования с изометрическими упражнениями, в которых было доказано, что давление кислорода (PO 2 ) в митохондриях низкое, но все же продемонстрировано увеличение окислительного стресса [26].Бейли и др. [21] также показали, что отток свободных радикалов из сокращающейся мышцы был связан со снижением внутриклеточного PO 2 , а не с условиями увеличения потока кислорода.
Это может происходить из-за неадекватной связи переноса электрона между комплексами I и III (рис. 1) [23, 24].По-видимому, комплекс I, а именно железо-серные кластеры, высвобождает активные формы кислорода — анион супероксида — только в направлении митохондриального матрикса, тогда как комплекс III — сайт окисления убихинола — высвобождает супероксид как в матрикс, так и за пределы внутренней мембраны [24]. , 25]. Несмотря на эту теоретическую привлекательность, существует мало прямых доказательств того, что производство митохондриальных супероксидных анионов увеличивается во время физических упражнений. Кроме того, есть исследования с изометрическими упражнениями, в которых было доказано, что давление кислорода (PO 2 ) в митохондриях низкое, но все же продемонстрировано увеличение окислительного стресса [26].Бейли и др. [21] также показали, что отток свободных радикалов из сокращающейся мышцы был связан со снижением внутриклеточного PO 2 , а не с условиями увеличения потока кислорода.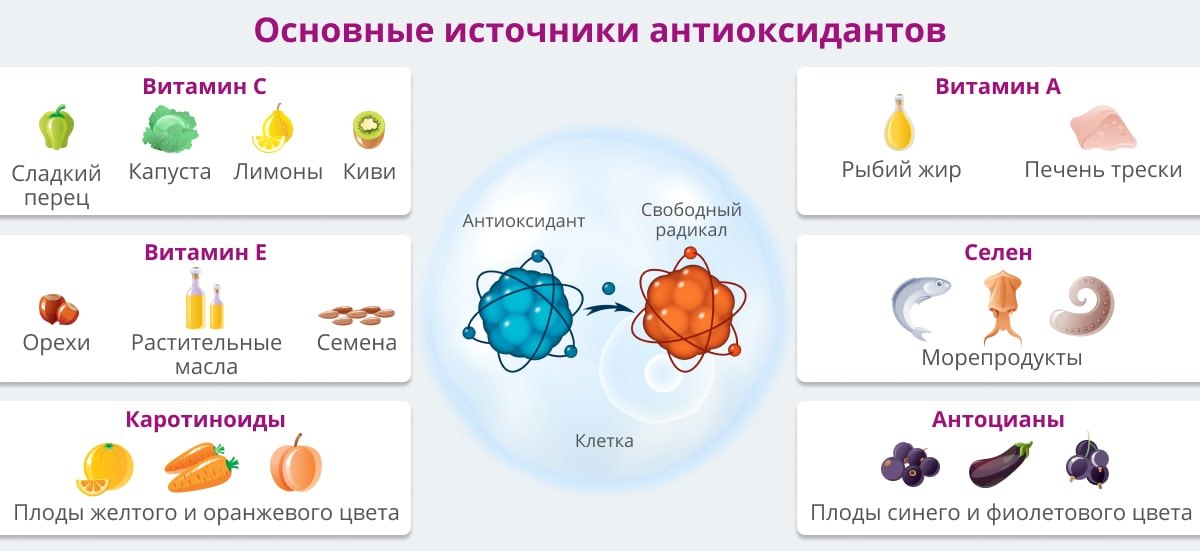 Эти исследования подтверждают тот факт, что маловероятно, что увеличение митохондриального потока кислорода является единственной или основной причиной увеличения выработки радикалов во время упражнений [23].
Эти исследования подтверждают тот факт, что маловероятно, что увеличение митохондриального потока кислорода является единственной или основной причиной увеличения выработки радикалов во время упражнений [23].
Реперфузия ишемии и активация эндотелиальной ксантиноксидазы
Известно, что фермент ксантиноксидаза участвует в патофизиологии синдрома ишемии-реперфузии и может привести к повреждению тканей, которое может произойти после изнурительной тренировки [27]. ].Следующие механизмы описывают этот процесс. Во время упражнений кровоток шунтируется от многих органов и тканей и перенаправляется на работающие мышцы; это ишемическое состояние запускает превращение фермента ксантиндегидрогеназы в ксантиноксидазу; когда упражнения прекращаются и ткани повторно оксигенируются, ксантиноксидаза производит супероксид (O2-) и H 2 O 2 в качестве побочных продуктов деградации гипоксантина в ксантин, а затем в мочевую кислоту [7], рис. 2.Хотя это было показано в нескольких исследованиях, необходимы дополнительные исследования, чтобы определить роль, которую эндотелиальная ксантиноксидаза играет в продукции АФК, вызванной физическими упражнениями, у людей.
Нейтрофилы и воспалительная реакция
В результате тренировки повреждение тканей или увеличение пула воспалительных клеток может привести к увеличению продукции реактивных частиц из немышечного источника. Когда нейтрофилы или другие фагоцитирующие клетки активируются, они выделяют АФК.Несмотря на то, что эта воспалительная реакция имеет решающее значение для удаления поврежденных белков и инфекций, АФК и другие оксиданты, высвобождаемые этими клетками, также могут вызывать вторичные повреждения, такие как перекисное окисление липидов. Упражнения могут вызвать повреждение мышц, сопровождающееся активацией нейтрофилов [28]. Бойюм и др. [29] продемонстрировали не только увеличение числа нейтрофилов после аэробных упражнений (езда на велосипеде в течение 65 мин при 75% VO2max), но и увеличение активности их дыхательных всплесков, измеряемое как хемилюминесценция.Увеличение количества нейтрофилов также было описано при коротких (менее 20 минут) упражнениях с отягощениями [30]. Это подтверждает тот факт, что количество нейтрофилов в плазме увеличивается даже тогда, когда потребление кислорода во время физической активности увеличивается лишь умеренно.
Это подтверждает тот факт, что количество нейтрофилов в плазме увеличивается даже тогда, когда потребление кислорода во время физической активности увеличивается лишь умеренно.
Структура НАДФН-оксидазы
Восстановленная форма комплекса никотинамидадениндинуклеотидфосфат (НАДФН) оксидазы была первоначально идентифицирована и охарактеризована в фагоцитах, где она играет важную роль в неспецифической защите хозяина от микробных организмов [31].Тем не менее, этот комплекс также обнаруживается во многих местах в скелетных мышцах, клеточном эндотелии и плазматической мембране [10]. НАДФН-оксидаза обычно находится в состоянии покоя, но когда она активируется, во время мышечного сокращения или когда ее задействуют для антимикробных и провоспалительных явлений, она может генерировать большие количества O2•-, которые могут быть преобразованы в H 2 O 2 под действием антиоксиданта. супероксиддисмутазы [31].
Самоокисление катехоламинов
Адреналин, норадреналин и дофамин часто вместе называют катехоламинами. При физической нагрузке отмечается повышение концентрации этих веществ в плазме [32]. Окисление катехоламинов может производить анион супероксида, H 2 O 2 и другие виды, не являющиеся производными кислорода, в сложной серии реакций. Это может привести к снижению концентрации клеточных антиоксидантов в крови, таких как глутатион, изменяя окислительно-восстановительный баланс [7, 10].
При физической нагрузке отмечается повышение концентрации этих веществ в плазме [32]. Окисление катехоламинов может производить анион супероксида, H 2 O 2 и другие виды, не являющиеся производными кислорода, в сложной серии реакций. Это может привести к снижению концентрации клеточных антиоксидантов в крови, таких как глутатион, изменяя окислительно-восстановительный баланс [7, 10].
Несмотря на необходимость лучшего понимания того, как взаимодействуют эти механизмы образования АФК, механизм, с помощью которого они влияют на функцию тренирующихся мышц, хорошо известен.В состоянии покоя низкая концентрация АФК в мышцах имеет решающее значение для выработки силы. Во время сократительной активности мышц увеличивается продукция АФК. С одной стороны, было показано, что производство реактивных частиц во время как аэробных, так и анаэробных неистощающих упражнений важно для адаптации мышечных волокон [33] — этот процесс будет обсуждаться в разделе 5. Тем не менее, во время напряженных упражнений производство АФК могут быть выше, чем буферная способность антиоксидантов мышц. По мере накопления АФК в сокращающихся мышцах окисление белков и липидов может вызывать, среди прочего, торможение производства силы, способствуя развитию острого утомления [10, 34]. Кроме того, такое преувеличенное повышение уровня АФК в ответ на интенсивные физические нагрузки также может приводить к окислительной модификации ДНК, ингибировать двигательную и бактерицидную активность нейтрофилов, снижать пролиферацию Т-лимфоцитов и В-лимфоцитов, ингибировать естественные клетки-киллеры, повреждать клеточную мембрану и другие клеточные соединения [35, 36].
По мере накопления АФК в сокращающихся мышцах окисление белков и липидов может вызывать, среди прочего, торможение производства силы, способствуя развитию острого утомления [10, 34]. Кроме того, такое преувеличенное повышение уровня АФК в ответ на интенсивные физические нагрузки также может приводить к окислительной модификации ДНК, ингибировать двигательную и бактерицидную активность нейтрофилов, снижать пролиферацию Т-лимфоцитов и В-лимфоцитов, ингибировать естественные клетки-киллеры, повреждать клеточную мембрану и другие клеточные соединения [35, 36].
4. Антиоксидантная система и упражнения
Из-за потенциальной роли реактивных частиц и свободных радикалов в повреждении липидов, белков и ДНК неудивительно, что в организме присутствует сеть механизмов антиоксидантной защиты. В целом, антиоксиданты часто являются восстанавливающими агентами, которые существуют как внутриклеточно, так и внеклеточно и обладают способностью реагировать со свободными радикалами и реактивными частицами, сводя к минимуму их действие и, таким образом, задерживая или предотвращая окислительный стресс [10].
Антиоксиданты могут синтезироваться in vivo и всасываться с пищей. Их можно разделить на две группы: ферментативные и неферментативные. К основным ферментным антиоксидантам относятся супероксиддисмутаза (СОД), глутатионпероксидаза (ГПХ) и каталаза (КАТ). Каждый из этих ферментов отвечает за восстановление разных АФК, и они расположены в разных клеточных компартментах. (1) СОД: существует 3 изоформы этого антиоксиданта, две из них присутствуют внутри клеток, а другая находится во внеклеточном пространстве.В частности, в клетках скелетных мышц самый высокий процент СОД (65–85%) находится в цитозоле, а оставшаяся часть (15–35%) присутствует в митохондриях мышц. СОД катализирует реакцию супероксидных радикалов с образованием кислорода и пероксидов водорода (H 2 O 2 ). (2) GPX: расположен как в цитозоле, так и в митохондриях клеток, он отвечает за удаление широкого спектра гидропероксидов — от сложных органических гидропероксидов до H 2 O 2 — таким образом, он может защищать мембранные липиды.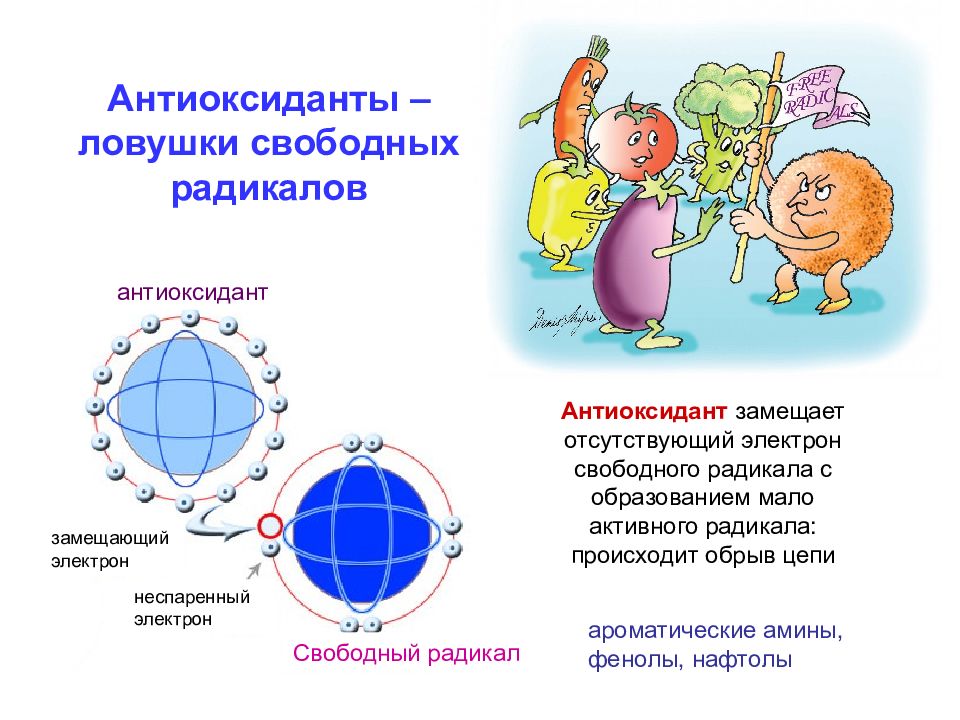 , белки и нуклеиновые кислоты от окисления.GPX также присутствует в мышечных клетках, но его активность варьируется в зависимости от типа мышечного волокна, при этом наибольшая активность присутствует в медленно сокращающихся мышечных волокнах (тип I), которые обладают более высокой окислительной способностью. (3) CAT: он широко распространен внутри клеток, и его основная функция заключается в разложении H 2 O 2 на H 2 O и O 2 . Тем не менее, он имеет более низкое сродство к H 2 O 2 по сравнению с GPX. Подобно последнему, CAT может быть обнаружен в более высокой концентрации в мышечных волокнах типа I (более подробную информацию об этих ферментах см. у Powers and Jackson [10]).
, белки и нуклеиновые кислоты от окисления.GPX также присутствует в мышечных клетках, но его активность варьируется в зависимости от типа мышечного волокна, при этом наибольшая активность присутствует в медленно сокращающихся мышечных волокнах (тип I), которые обладают более высокой окислительной способностью. (3) CAT: он широко распространен внутри клеток, и его основная функция заключается в разложении H 2 O 2 на H 2 O и O 2 . Тем не менее, он имеет более низкое сродство к H 2 O 2 по сравнению с GPX. Подобно последнему, CAT может быть обнаружен в более высокой концентрации в мышечных волокнах типа I (более подробную информацию об этих ферментах см. у Powers and Jackson [10]).
Группа неферментативных антиоксидантов включает глутатион, витамин С, витамин Е, каротиноиды, мочевую кислоту и другие. Подобно ферментативным антиоксидантам, они присутствуют в разных клеточных компартментах и проявляют различные антиоксидантные свойства, которые максимизируют их эффективность [11]. Ниже приведены более подробные сведения о неферментативных антиоксидантах — глутатионе, витамине С и витамине Е. глицин.Эти три аминокислоты можно получить из пищи. Все типы клеток способны синтезировать GSH. Этот процесс синтеза происходит посредством двух последовательных реакций и требует действия двух ферментов: для первой реакции γ -глутамилцистеина и синтетазы GSH для последующей реакции (рис. 3). Уровни продуцируемого GSH действуют как обратная связь для контроля реакции 1 (см. Рисунок 3), это помогает поддерживать его адекватную продукцию [37]. GSH также может быть получен непосредственно из пищи, но только небольшое количество интактного трипептида всасывается в кишечнике; большинство расщепляется на аминокислоты, которые затем используются в цикле синтеза.Таким образом, здоровое питание со сбалансированным потреблением белка необходимо для адекватного гомеостаза GSH в организме [38, 39].
Ниже приведены более подробные сведения о неферментативных антиоксидантах — глутатионе, витамине С и витамине Е. глицин.Эти три аминокислоты можно получить из пищи. Все типы клеток способны синтезировать GSH. Этот процесс синтеза происходит посредством двух последовательных реакций и требует действия двух ферментов: для первой реакции γ -глутамилцистеина и синтетазы GSH для последующей реакции (рис. 3). Уровни продуцируемого GSH действуют как обратная связь для контроля реакции 1 (см. Рисунок 3), это помогает поддерживать его адекватную продукцию [37]. GSH также может быть получен непосредственно из пищи, но только небольшое количество интактного трипептида всасывается в кишечнике; большинство расщепляется на аминокислоты, которые затем используются в цикле синтеза.Таким образом, здоровое питание со сбалансированным потреблением белка необходимо для адекватного гомеостаза GSH в организме [38, 39].
GSH выполняет различные важные функции в организме. Среди этих функций его главная антиоксидантная роль. Он эффективно удаляет АФК и свободные радикалы, предотвращая усиление процесса окислительного стресса. В этих реакциях восстановленный GSH окисляется ферментом глутатионпероксидазой с образованием глутатиондисульфида (GSSG). Обратите внимание, что GSSG образован двумя молекулами GSH, связанными дисульфидной связью за счет окисления тиоловых (SH) групп.После окисления GSSG может быть восстановлен до исходной формы GSH с помощью фермента GSSG-редуктазы и никотинамидадениндинуклеотидфосфата (NADPH). Тем не менее, при высоком уровне окислительного стресса происходит истощение НАДФН и внутриклеточное накопление GSSG. Этот избыток GSSG может либо экспортироваться из клетки, либо образовывать смешанный дисульфид. Несмотря на то, что происходит с окисленным глутатионом, истощение клеточного GSH может наблюдаться, когда происходит интенсивный процесс окислительного стресса [40].
Он эффективно удаляет АФК и свободные радикалы, предотвращая усиление процесса окислительного стресса. В этих реакциях восстановленный GSH окисляется ферментом глутатионпероксидазой с образованием глутатиондисульфида (GSSG). Обратите внимание, что GSSG образован двумя молекулами GSH, связанными дисульфидной связью за счет окисления тиоловых (SH) групп.После окисления GSSG может быть восстановлен до исходной формы GSH с помощью фермента GSSG-редуктазы и никотинамидадениндинуклеотидфосфата (NADPH). Тем не менее, при высоком уровне окислительного стресса происходит истощение НАДФН и внутриклеточное накопление GSSG. Этот избыток GSSG может либо экспортироваться из клетки, либо образовывать смешанный дисульфид. Несмотря на то, что происходит с окисленным глутатионом, истощение клеточного GSH может наблюдаться, когда происходит интенсивный процесс окислительного стресса [40].
Измерение уровня GSH или его окисленной формы (GSSG) в плазме является широко распространенным методом выявления окислительного стресса и может быть представлено как окислительно-восстановительный потенциал, концентрация GSH или GSSG или отношение GSH/GSSG. Это не только хороший показатель системного окислительного статуса, но и полезный индикатор риска заболевания [41], и поэтому он использовался в различных исследованиях для определения образования свободных радикалов во время физических упражнений [19, 42–44]. Однако в этих исследованиях представлены некоторые расходящиеся результаты, которые, возможно, можно объяснить разницей в физической подготовке участников, протоколе упражнений и методе определения концентрации глутатиона.
Это не только хороший показатель системного окислительного статуса, но и полезный индикатор риска заболевания [41], и поэтому он использовался в различных исследованиях для определения образования свободных радикалов во время физических упражнений [19, 42–44]. Однако в этих исследованиях представлены некоторые расходящиеся результаты, которые, возможно, можно объяснить разницей в физической подготовке участников, протоколе упражнений и методе определения концентрации глутатиона.
Витамин С
Это водорастворимый витамин, относящийся как к аскорбиновой кислоте, так и к дегидроаскорбиновой кислоте (ДГК). Аскорбиновая кислота является основной формой витамина, найденного в естественных условиях . Этот витамин, также называемый аскорбатом, содержится в относительно больших количествах в различных тканях по всему телу. Было ясно показано, что аскорбат играет важную роль в биосинтезе соединительной ткани, а его дефицит приводит к цинге, заболеванию, которое приводит к ухудшению выработки коллагена и приводит к хрупкости кровеносных сосудов и нарушению заживления повреждений. Это заболевание обратимо, как только человек возобновляет прием аскорбата. Витамин С также является сильным восстановителем благодаря своей способности отдавать электроны и обладает важными антиоксидантными свойствами [45]. Он может инактивировать различные реактивные виды, сводя к минимуму повреждение тканей организма.
Это заболевание обратимо, как только человек возобновляет прием аскорбата. Витамин С также является сильным восстановителем благодаря своей способности отдавать электроны и обладает важными антиоксидантными свойствами [45]. Он может инактивировать различные реактивные виды, сводя к минимуму повреждение тканей организма.
Во время реакций окисления теряется лишь небольшое количество аскорбата, потому что после окисления он может снова восстановиться до аскорбиновой кислоты восстановителями, такими как глутатион, никотинамидадениндинуклеотид (НАДН) и НАДФН.Точно так же известно, что витамин С регенерирует другие антиоксиданты, такие как витамин Е и глутатион, обратно в их восстанавливающее состояние; таким образом, поддерживая сбалансированную сеть антиоксидантов [46].
При приеме внутрь витамин С всасывается в кишечнике либо путем активного транспорта, если доступность витамина низкая, либо путем простой диффузии, если он присутствует в высоких концентрациях. При попадании в организм в избытке он разлагается в кишечнике и может вызвать диарею и дискомфорт в кишечнике. Помимо механизмов абсорбции и деградации, происходящих в кишечнике, почки также несут ответственность за сохранение или удаление неметаболизированной аскорбиновой кислоты, чтобы помочь поддерживать баланс аскорбиновой кислоты в организме [45].
Помимо механизмов абсорбции и деградации, происходящих в кишечнике, почки также несут ответственность за сохранение или удаление неметаболизированной аскорбиновой кислоты, чтобы помочь поддерживать баланс аскорбиновой кислоты в организме [45].
Витамин Е
Является жирорастворимым витамином, также называемым α -токоферолом. Молекулы витамина Е различаются по структуре. Различные формы этого витамина значительно различаются по своим метаболическим функциям и биодоступности.У людей более 90% витамина Е, встречающегося в организме, представляет собой α -токоферол. Однако не все формы α -токоферола сохраняются в плазме. α -токоферол, встречающийся в природе в пищевых продуктах вместе с синтетическими формами α -токоферола (изомеры RRR-, RSR-, RRS- и RSS- α -токоферол), может поддерживаться в плазма и ткани человека. При приеме внутрь добавок α-токоферола предпочтительна форма RRR-α-токоферола [45, 47].
Было показано, что этот витамин оказывает благотворное влияние на некоторые заболевания, он также связан со снижением риска сердечно-сосудистых заболеваний и может помочь замедлить развитие дегенеративных заболеваний, таких как атеросклероз. Напротив, у пациентов, страдающих нарушением всасывания жира в кишечнике, дефицит α-токоферола был связан с дегенерацией нейронов [48]. Тем не менее, недавнее обзорное исследование [49], в котором тщательно проанализировано более 47 исследований антиоксидантных добавок по смертности от всех причин, неожиданно показало негативное влияние некоторых добавок.Витамин Е, например, был одним из антиоксидантов, который был связан с увеличением смертности, в то время как витамин С не проявлял подобного эффекта. Трудно определить конкретные биохимические и физиологические механизмы, которые могли привести к такому результату. Поэтому необходимы дополнительные исследования, чтобы ответить на вопросы, которые возникают в результате таких исследований. Кроме того, эти результаты, касающиеся синтетических антиоксидантов, не следует переносить на влияние фруктов и овощей на здоровье человека.
Напротив, у пациентов, страдающих нарушением всасывания жира в кишечнике, дефицит α-токоферола был связан с дегенерацией нейронов [48]. Тем не менее, недавнее обзорное исследование [49], в котором тщательно проанализировано более 47 исследований антиоксидантных добавок по смертности от всех причин, неожиданно показало негативное влияние некоторых добавок.Витамин Е, например, был одним из антиоксидантов, который был связан с увеличением смертности, в то время как витамин С не проявлял подобного эффекта. Трудно определить конкретные биохимические и физиологические механизмы, которые могли привести к такому результату. Поэтому необходимы дополнительные исследования, чтобы ответить на вопросы, которые возникают в результате таких исследований. Кроме того, эти результаты, касающиеся синтетических антиоксидантов, не следует переносить на влияние фруктов и овощей на здоровье человека.
Подобно витамину С, витамин Е обладает важными антиоксидантными свойствами. Благодаря своей способности улавливать АФК и свободные радикалы, особенно пероксильный радикал (ROO•), он выполняет важную функцию защиты клеточных мембран и липопротеинов плазмы от перекисного окисления липидов. Это возможно, поскольку витамин Е обладает большим сродством к восстановлению пероксильных радикалов, предотвращая их взаимодействие с мембранными фосфолипидами или липопротеинами [45]. Действительно, витамин Е можно считать одним из основных ингибиторов перекисного окисления липидов in vivo [7].После окисления он может быть регенерирован обратно в восстановленное состояние с помощью витамина С, как упоминалось ранее. Тем не менее, исследования показывают, что повышенные уровни радикалов α -токоферола, которые не преобразуются обратно в восстановленную форму, могут действовать как прооксиданты, сами по себе инициируя процессы окислительного стресса. Кроме того, среда, в которой отсутствуют другие антиоксиданты или в которой наблюдается высокий уровень окислительного стресса, будет способствовать тому, что витамин Е будет действовать как прооксидант [46].
Это возможно, поскольку витамин Е обладает большим сродством к восстановлению пероксильных радикалов, предотвращая их взаимодействие с мембранными фосфолипидами или липопротеинами [45]. Действительно, витамин Е можно считать одним из основных ингибиторов перекисного окисления липидов in vivo [7].После окисления он может быть регенерирован обратно в восстановленное состояние с помощью витамина С, как упоминалось ранее. Тем не менее, исследования показывают, что повышенные уровни радикалов α -токоферола, которые не преобразуются обратно в восстановленную форму, могут действовать как прооксиданты, сами по себе инициируя процессы окислительного стресса. Кроме того, среда, в которой отсутствуют другие антиоксиданты или в которой наблюдается высокий уровень окислительного стресса, будет способствовать тому, что витамин Е будет действовать как прооксидант [46].
Доказательства того, что физические упражнения приводят к увеличению количества свободных радикалов и реактивных частиц, побудили исследователей проанализировать эффективность пищевых добавок с антиоксидантами, чтобы ослабить окислительный стресс в мышцах и, таким образом, улучшить мышечную работоспособность и иммунную функцию. В качестве примера можно упомянуть добавки с витамином Е. Как упоминалось в начале этого раздела, было обнаружено, что витамин Е защищает клеточные мембраны от перекисного окисления липидов. Следовательно, логично предположить, что этот витамин может защищать мышечные клетки от повреждений, вызванных физической нагрузкой. Ранние исследования, анализирующие влияние добавок витамина Е и физических упражнений, изучали его влияние на работоспособность. Однако в большинстве исследований не сообщается о пользе витамина Е ни для мышечной силы, ни для выносливости [50].Кроме того, было высказано предположение, что добавка витамина Е может оказывать защитное действие против окислительного стресса, вызванного сокращением мышц, который может возникнуть после интенсивной тренировки. Это обоснование основано на знании того, что этот витамин может стабилизировать мышечные мембраны, взаимодействуя с его фосфолипидами, которые, таким образом, обеспечивают некоторую защиту от увеличения окислительного стресса или повреждения мышц, наблюдаемого после определенных видов упражнений [50].
В качестве примера можно упомянуть добавки с витамином Е. Как упоминалось в начале этого раздела, было обнаружено, что витамин Е защищает клеточные мембраны от перекисного окисления липидов. Следовательно, логично предположить, что этот витамин может защищать мышечные клетки от повреждений, вызванных физической нагрузкой. Ранние исследования, анализирующие влияние добавок витамина Е и физических упражнений, изучали его влияние на работоспособность. Однако в большинстве исследований не сообщается о пользе витамина Е ни для мышечной силы, ни для выносливости [50].Кроме того, было высказано предположение, что добавка витамина Е может оказывать защитное действие против окислительного стресса, вызванного сокращением мышц, который может возникнуть после интенсивной тренировки. Это обоснование основано на знании того, что этот витамин может стабилизировать мышечные мембраны, взаимодействуя с его фосфолипидами, которые, таким образом, обеспечивают некоторую защиту от увеличения окислительного стресса или повреждения мышц, наблюдаемого после определенных видов упражнений [50]. Тем не менее, результаты различных исследований были довольно противоречивыми, и подробное обсуждение этих вопросов не входит в рамки данного обзора, для получения дополнительной информации см. обзор Jackson et al.[51].
Тем не менее, результаты различных исследований были довольно противоречивыми, и подробное обсуждение этих вопросов не входит в рамки данного обзора, для получения дополнительной информации см. обзор Jackson et al.[51].
Интересно, что недавняя работа показала, что увеличение реактивных частиц во время упражнений приводит к благоприятной адаптации, вызванной физическими упражнениями. В соответствующей литературе хорошо задокументировано, что как аэробные, так и анаэробные тренировки вызывают повышение активности антиоксидантных ферментов в различных тканях [33, 52, 53]. Это процесс адаптации, который происходит потому, что свободные радикалы, образующиеся во время мышечного сокращения, действуют как сигнальные молекулы. Это стимулирует экспрессию генов и, следовательно, увеличивает выработку антиоксидантных ферментов и модулирует другие пути защиты от окислительного стресса, такие как усиление активности ферментов репарации ДНК в скелетных мышцах [33, 54].Это укрепляет систему антиоксидантной сети организма, что, следовательно, минимизирует процесс окислительного стресса [33]. Эти стимулы, связанные с усиленной антиоксидантной защитой, возникают не только в мышцах, но и системно, поэтому жизненно важные органы, такие как печень и мозг, также проходят через эту полезную адаптацию [55].
Эти стимулы, связанные с усиленной антиоксидантной защитой, возникают не только в мышцах, но и системно, поэтому жизненно важные органы, такие как печень и мозг, также проходят через эту полезную адаптацию [55].
Этот процесс адаптации, возникающий в результате регулярных физических упражнений, можно объяснить с помощью теории гормезиса. Эта теория постулирует, что химические вещества и токсические вещества могут иметь стимулирующий эффект в малых дозах и ингибирующий эффект в высоких дозах.Это означает, что они могут давать положительные ответы, когда присутствуют в небольшом количестве [56]. В связи с этим тип и продолжительность тренировок являются ключевыми для значительного повышения уровня эндогенных антиоксидантов, при этом более эффективными являются длительные высокоинтенсивные тренировки на выносливость [57]. Нисс и др. [58] сообщили, что у тренированных людей было меньше повреждений ДНК после изнурительной тренировки по сравнению с нетренированными мужчинами. Другие доказательства этой адаптации обучения были представлены Miyazaki et al. [59], которые показали, что выработка свободных радикалов снижалась после 12 недель тренировок на выносливость.В частности, они сообщили о снижении продукции супероксидных анионов нейтрофилами и ослаблении процесса перекисного окисления липидов. Верно и обратное, поскольку малоподвижный образ жизни не только снижает различные физиологические функции, но и снижает защитные механизмы организма от окислительного стресса. Следовательно, может увеличиться возникновение окислительного стресса, связанного с такими заболеваниями, как рак, атеросклероз, сердечно-сосудистые и нейродегенеративные заболевания [55].
[59], которые показали, что выработка свободных радикалов снижалась после 12 недель тренировок на выносливость.В частности, они сообщили о снижении продукции супероксидных анионов нейтрофилами и ослаблении процесса перекисного окисления липидов. Верно и обратное, поскольку малоподвижный образ жизни не только снижает различные физиологические функции, но и снижает защитные механизмы организма от окислительного стресса. Следовательно, может увеличиться возникновение окислительного стресса, связанного с такими заболеваниями, как рак, атеросклероз, сердечно-сосудистые и нейродегенеративные заболевания [55].
5.Играет ли АФК важную роль в реакции адаптации, вызванной физической нагрузкой?
Как упоминалось ранее, окислительный стресс часто используется для обозначения состояния, при котором накопление реактивных частиц оказывает повреждающее действие на многие ткани и органы [7]. Тем не менее, новые данные показали, что окислительный стресс играет решающую роль в гомеостазе мышц и метаболизме скелетных мышц при физической нагрузке.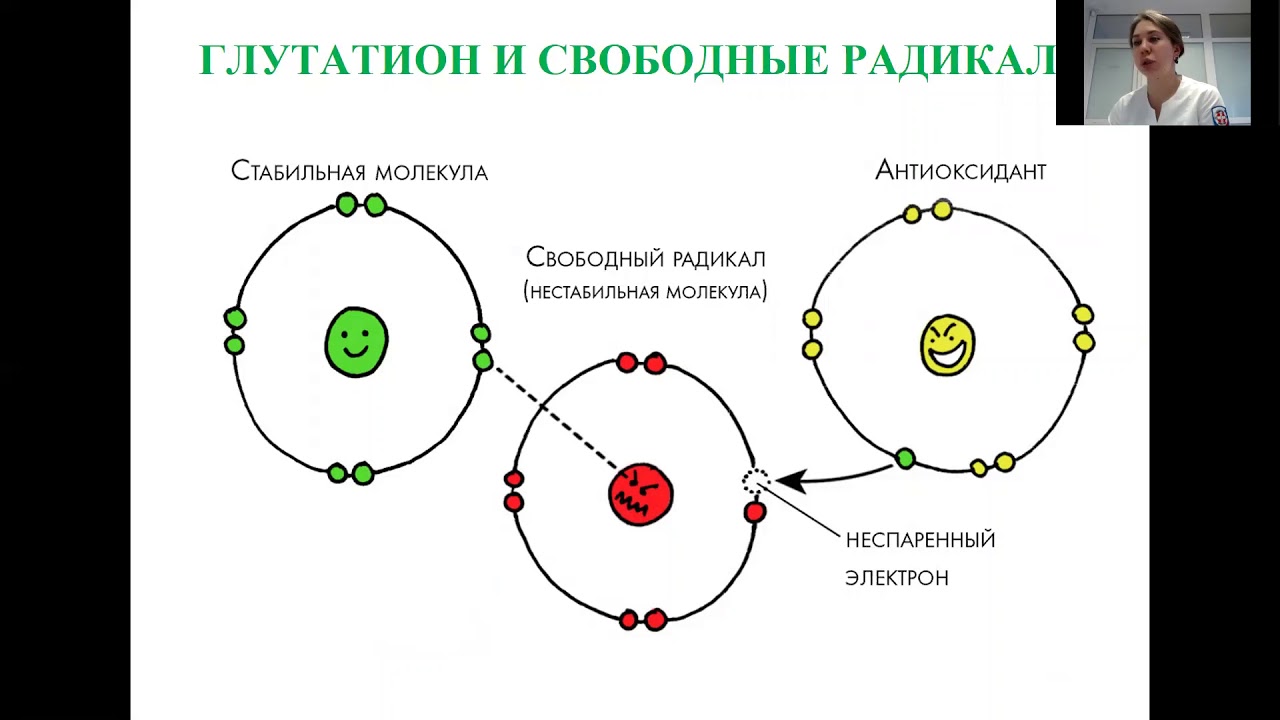 Это говорит о том, что реактивные виды являются не просто повреждающими агентами, вызывающими случайные разрушения клеточных структур и функций, но также — по крайней мере, в пределах физиологической концентрации — полезными сигнальными молекулами, которые регулируют рост, пролиферацию, дифференцировку и могут быть ответственны за некоторые адаптации в клетках. в тканях, подвергаемых тренировке, и в тканях, не подвергающихся тренировке, таких как мозг [55, 60].Следовательно, физические упражнения вызывают резкое увеличение продукции реактивных частиц в скелетных мышцах, которые могут работать как внутриклеточные сигнальные молекулы [61, 62]. Но как реактивные виды действуют как физиологические сигнальные молекулы в ремоделирующей ткани, вызванной физическими упражнениями, и кто является наиболее важными игроками в этой игре?
Это говорит о том, что реактивные виды являются не просто повреждающими агентами, вызывающими случайные разрушения клеточных структур и функций, но также — по крайней мере, в пределах физиологической концентрации — полезными сигнальными молекулами, которые регулируют рост, пролиферацию, дифференцировку и могут быть ответственны за некоторые адаптации в клетках. в тканях, подвергаемых тренировке, и в тканях, не подвергающихся тренировке, таких как мозг [55, 60].Следовательно, физические упражнения вызывают резкое увеличение продукции реактивных частиц в скелетных мышцах, которые могут работать как внутриклеточные сигнальные молекулы [61, 62]. Но как реактивные виды действуют как физиологические сигнальные молекулы в ремоделирующей ткани, вызванной физическими упражнениями, и кто является наиболее важными игроками в этой игре?
5.1. Ангиогенез
Ангиогенез – это процесс, посредством которого активированные эндотелиальные клетки отходят от существующего капилляра [63]. Рост новых кровеносных сосудов является важным естественным процессом, необходимым для заживления ран и восстановления кровотока в тканях после травмы или инсульта [64]. Известно, что тренировка на выносливость вызывает реакции ангиогенеза в мышечной ткани [63]. Для спортсменов на выносливость эта адаптация очень важна из-за повышенной потребности в снабжении кислородом всей мышечной массы. Процесс ангиогенеза контролируется рядом медиаторов, высвобождаемых в тканях, окружающих мелкие сосуды [64]. Физические упражнения могут активировать этот процесс, способствуя высвобождению некоторых медиаторов. Значительным изменениям (физиологическим или физиопатологическим) механических сил, возникающих 90–111 in vivo 90–112, нельзя противопоставить острую регуляцию диаметра стенки сосуда, что приводит к фенотипической модуляции эндотелиальных и сосудистых клеток, вызывая структурные модификации артериальной стенки.Таким образом, ремоделирование сосудов является фундаментальной основой нормального роста сосудов и адаптации к физической нагрузке.
Известно, что тренировка на выносливость вызывает реакции ангиогенеза в мышечной ткани [63]. Для спортсменов на выносливость эта адаптация очень важна из-за повышенной потребности в снабжении кислородом всей мышечной массы. Процесс ангиогенеза контролируется рядом медиаторов, высвобождаемых в тканях, окружающих мелкие сосуды [64]. Физические упражнения могут активировать этот процесс, способствуя высвобождению некоторых медиаторов. Значительным изменениям (физиологическим или физиопатологическим) механических сил, возникающих 90–111 in vivo 90–112, нельзя противопоставить острую регуляцию диаметра стенки сосуда, что приводит к фенотипической модуляции эндотелиальных и сосудистых клеток, вызывая структурные модификации артериальной стенки.Таким образом, ремоделирование сосудов является фундаментальной основой нормального роста сосудов и адаптации к физической нагрузке.
Недавно роль АФК в индуцированном высоким потоком ремоделировании сосудов была продемонстрирована на мышиной модели артерии и яремной вены (АВФ), где продукция АФК увеличивалась в артериях, подверженных хроническому высокому потоку, как через 1, так и через 3 недели после открытия АВФ [65]. НАДФН-оксидаза идентифицирована как основной генератор АФК, индуцированных сдвиговым напряжением, в сосудистой стенке АВФ. Генерация АФК и NO в результате активации эндотелиальной оксид азотсинтазы (eNOS) приводила к активации матриксных металлопротеиназ (ММП) при АВФ и модулированию вызванного потоком расширения сосудов.ММП в совокупности расщепляют большинство, если не все, составляющие внеклеточного матрикса. Наблюдения, проведенные на мышах p47phox -/- и мышах eNOS -/- , также предоставили прямые доказательства того, что эндогенные АФК и NO модулируют не только активность ММР, но и их продукцию в артериях, подверженных повышенному кровотоку. Lehoux [66] показал, что долговременная структурная адаптация к измененному кровотоку опосредуется АФК. Кроме того, Lehoux и соавт. [67] продемонстрировали, что ММР-9, индуцированный высоким внутрипросветным давлением в сонных артериях, способствует увеличению растяжимости сосудов.Этот фактор указывает на роль ферментов, разрушающих матрикс, на ранних стадиях ремоделирования сосудов.
НАДФН-оксидаза идентифицирована как основной генератор АФК, индуцированных сдвиговым напряжением, в сосудистой стенке АВФ. Генерация АФК и NO в результате активации эндотелиальной оксид азотсинтазы (eNOS) приводила к активации матриксных металлопротеиназ (ММП) при АВФ и модулированию вызванного потоком расширения сосудов.ММП в совокупности расщепляют большинство, если не все, составляющие внеклеточного матрикса. Наблюдения, проведенные на мышах p47phox -/- и мышах eNOS -/- , также предоставили прямые доказательства того, что эндогенные АФК и NO модулируют не только активность ММР, но и их продукцию в артериях, подверженных повышенному кровотоку. Lehoux [66] показал, что долговременная структурная адаптация к измененному кровотоку опосредуется АФК. Кроме того, Lehoux и соавт. [67] продемонстрировали, что ММР-9, индуцированный высоким внутрипросветным давлением в сонных артериях, способствует увеличению растяжимости сосудов.Этот фактор указывает на роль ферментов, разрушающих матрикс, на ранних стадиях ремоделирования сосудов.
Как хорошо известно, ключом к ангиогенезу является фактор роста эндотелия сосудов (VEGF), но реальная связь между физической нагрузкой и ангиогенезом до конца не изучена. Первоначальные исследования по этой теме проводились на скелетных мышцах. В скелетных мышцах крыс происходит активация VEGF после хронической мышечной стимуляции или однократного бега на беговой дорожке с умеренной интенсивностью [68].В случае бега на беговой дорожке уровень мРНК VEGF повышался в активных мышцах крыс примерно в 2–4 раза к концу тренировки и оставался повышенным в течение следующих 4 часов. Через восемь часов после тренировки уровни мРНК VEGF вернулись к таковым в покоящихся мышцах.
Согласно Gustafsson et al. [69], активация мРНК VEGF также происходит во время физических упражнений у здоровых людей. Точно так же физические упражнения активируют другие элементы, важные для процесса ангиогенеза, например, мРНК рецепторов VEGF 1 и 2 (VEGFR1 и VEGFR2 соответственно.) увеличиваются после мышечной активности [70]. Таким образом, кажется, что аэробные упражнения могут усиливать ангиогенный каскад.
Таким образом, кажется, что аэробные упражнения могут усиливать ангиогенный каскад.
Другим важным и недавно исследованным игроком в процессе ангиогенеза, вызванном физической нагрузкой, является PGC-1 α (коактиватор рецептора-c, активируемый пероксисомой-пролифератором-1 α ). Арани и др. [71] показали, что PGC-1 α является мощным датчиком и регулятором метаболизма, индуцируемым недостатком питательных веществ и кислорода. Кроме того, PGC-1 α мощно регулирует экспрессию VEGF и ангиогенез в культивируемых мышечных клетках и скелетных мышцах in vivo .Упражнения могут вызвать временную ишемию мышечной ткани [72]. Ишемия приводит к серьезным метаболическим проблемам с потенциально катастрофическими последствиями. Например, у мышей PGC-1 α -/- наблюдалась поразительная неспособность восстановить нормальный кровоток в конечности после ишемического инсульта, тогда как трансгенная экспрессия PGC-1 α в скелетных мышцах является защитным.
Вест и др. [73] описали новый механизм ангиогенеза, который не зависит от запускаемой гипоксией экспрессии VEGF.Продукты перекисного окисления липидов образуются как следствие окислительного стресса и распознаются толл-подобными рецепторами (TLR), стимулируя ангиогенез in vivo , тем самым способствуя ускорению заживления ран и восстановлению тканей. Капиллярность в активных скелетных мышцах значительно увеличивается при тренировках на выносливость, и, согласно Bloor [63], любое увеличение капиллярности мышц важно для улучшения свойств обмена крови в тканях. Это связано с тем, что большая капиллярная сеть (1) увеличивает площадь поверхности для диффузии, (2) сокращает среднюю длину пути диффузии внутри мышцы и (3) увеличивает продолжительность диффузионного обмена между кровью и тканью.Следовательно, персистенция вновь образованной сосудистой сети в воспалительном контексте приводит к сохранению ремоделирования ткани. Необходимы дополнительные исследования, чтобы определить взаимосвязь иммунных клеток и других молекул и рецепторов, таких как VEGF, PGC1- α и TLR2, во время процесса новообразования сосудов в ответ на физическую нагрузку.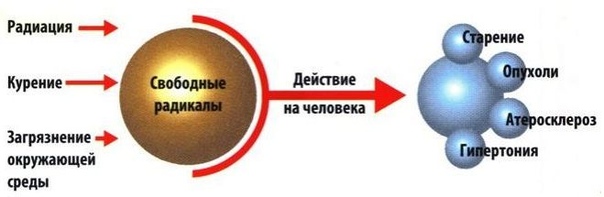 Это имело бы значение для создания стратегий обучения для оптимального восстановления тканей.
Это имело бы значение для создания стратегий обучения для оптимального восстановления тканей.
5.2. Митохондриальный биогенез
Регулярные аэробные тренировки вызывают адаптацию в скелетных мышцах, называемую митохондриальным биогенезом [74], то есть увеличение количества и размера митохондрий [75].Механизм этой адаптации не ясен, но недавние исследования предполагают возможную роль АФК в этом процессе. Хотя значительное внимание уделялось повреждению, вызванному продукцией АФК, также известно, что АФК могут активировать сигнальные пути, участвующие в фенотипической адаптации. Исследования показывают, что АФК вызывают разветвление и удлинение митохондриальной сети. Пеше и др. [76] показали, что количество копий мтДНК увеличивалось с повышением уровня АФК в стареющих скелетных мышцах, и это увеличение мтДНК сопровождалось индукцией митохондриальной массы.Этот ответ, по-видимому, опосредован PGC-1 α и ядерным респираторным фактором 1 (NRF-1), поскольку экспрессия обоих увеличилась после экзогенной обработки АФК [77]. Недавно Иррчер и соавт. [78] продемонстрировали, что АФК могут приводить к увеличению активности и экспрессии промотора PGC-1 α как посредством зависимого от аденозинмонофосфат-активируемой протеинкиназы (AMPK), так и независимого от AMPK пути. Эти пути, скорее всего, частично объясняют усиление митохондриального биогенеза, наблюдаемое в присутствии АФК.
5.3. Гипертрофия скелетных мышц
В литературе предполагается, что реактивные виды играют важную роль в регуляции клеточной передачи сигналов и изменений в экспрессии генов [79, 80], способствуя, среди прочего, контролю размера скелетных мышц. Данные, полученные in vitro , показывают, что АФК способны влиять на эффективность дифференцировки мышечной ткани и успешную дифференцировку миобластов сателлитного происхождения (клеток-предшественников, образующих мышечные клетки) в функционирующие.Интегрированные мышечные трубки являются фундаментальной предпосылкой для регенерации мышц, процесса восстановления, который имеет первостепенное значение для поддержания мышечной функции [61].
АФК модулируют передачу сигналов различных факторов роста посредством окислительно-восстановительной регуляции. Однако роль АФК в передаче сигналов инсулиноподобного фактора роста-1 (ИФР-1) до конца не изучена. IGF-1 — это гормон с молекулярной структурой, сходной с инсулином. Он играет очень важную роль в пролиферации, дифференцировке и выживании клеток, выполняя различные тканеспецифические функции [81].IGF-1 индуцирует гипертрофию миоцитов (также известных как мышечные клетки), особенно в скелетных мышцах [82]. In vivo мышечно-специфические трансгенные мыши со сверхэкспрессией IGF-1 демонстрируют повышенную мышечную массу, а мыши со специфическим отсутствием рецептора IGF-1 (IGF-IR) в мышцах имеют меньшие мышцы и уменьшенное количество мышечных волокон [83]. Hadayaningsih и коллеги [84] недавно исследовали, регулируют ли АФК передачу сигналов и биологическое действие IGF-1 в миоцитах C2C12 у мышей. Они обнаружили, что IGF-1 индуцирует ROS в миоцитах C2C12; в то время как обработка H 2 O 2 значительно усиливала индуцированное IGF-I фосфорилирование рецептора IGF-1 (IGF-1R), фосфорилирование заметно ослаблялось при обработке клеток антиоксидантами.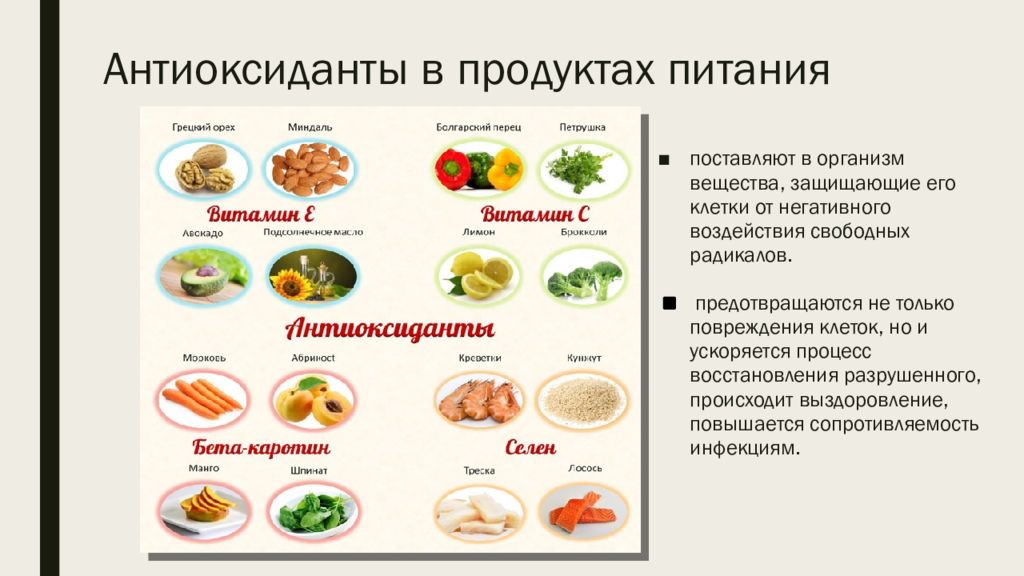 Нисходящий сигнальный путь Akt/mTOR-p70S6K (хорошо известный гипертрофический путь) впоследствии подавлялся, и, кроме того, блокирование Nox4, который, как сообщается, продуцирует АФК в передаче сигналов инсулина, ослабляло индуцированное IGF-1 фосфорилирование IGF-1R, что указывает на что Nox4 участвует в регуляции передачи сигналов IGF-1. Важно отметить, что антиоксидантная обработка ингибировала гипертрофию миоцитов, индуцированную IGF-1, демонстрируя, что АФК необходимы для индуцирования гипертрофии миоцитов IGF-1 in vitro .Эти результаты показывают, что реактивные виды играют важную роль в передаче сигналов и биологическом действии IGF-1 в миоцитах C2C12, ясно показывая, что IGF-1 является важным регулятором гипертрофии миоцитов. Поскольку силовые упражнения стимулируют гипертрофию скелетных мышц и локальную экспрессию ИФР-1, это предполагает участие ИФР-1 в гипертрофии миоцитов, вызванной физической нагрузкой.
Нисходящий сигнальный путь Akt/mTOR-p70S6K (хорошо известный гипертрофический путь) впоследствии подавлялся, и, кроме того, блокирование Nox4, который, как сообщается, продуцирует АФК в передаче сигналов инсулина, ослабляло индуцированное IGF-1 фосфорилирование IGF-1R, что указывает на что Nox4 участвует в регуляции передачи сигналов IGF-1. Важно отметить, что антиоксидантная обработка ингибировала гипертрофию миоцитов, индуцированную IGF-1, демонстрируя, что АФК необходимы для индуцирования гипертрофии миоцитов IGF-1 in vitro .Эти результаты показывают, что реактивные виды играют важную роль в передаче сигналов и биологическом действии IGF-1 в миоцитах C2C12, ясно показывая, что IGF-1 является важным регулятором гипертрофии миоцитов. Поскольку силовые упражнения стимулируют гипертрофию скелетных мышц и локальную экспрессию ИФР-1, это предполагает участие ИФР-1 в гипертрофии миоцитов, вызванной физической нагрузкой.
Еще одна молекула, играющая важную роль в гипертрофическом ответе на определенные упражнения, — это интерлейкин-6 (ИЛ-6). Недавняя научная литература идентифицирует IL-6 как важный регулятор опосредованного сателлитными клетками (мышечными стволовыми клетками) гипертрофического роста мышц. IL-6 локально и временно продуцируется растущими миофибриллами и ассоциированными сателлитными клетками, а генетическая потеря IL-6 притупляет мышечную гипертрофию in vivo [85]. Космиду и др. [86] показали, что АФК стимулируют высвобождение IL-6 из скелетных мышечных трубок, таким образом, идентифицируя новый клеточный источник, который может быть ответственным за избыточное количество цитокинов, чувствительных к воспалению, вырабатываемых в моделях с напряженными упражнениями.Их результаты также предполагают, что стимулированное АФК увеличение высвобождения IL-6 зависит от транскрипции и включает активацию p38 и NF𝜅-B. Хотя p38-MAPK появляется как ключевая молекула, управляющая последовательными событиями в миогенном пути, многие детали p38-MAPK-индуцированного миогенеза еще предстоит выяснить. Еще предстоит определить, регулируют ли различные члены семейства p38 MAPK экспрессию отдельных субнаборов генов на разных стадиях дифференцировки и обладают ли они индуцирующей или репрессирующей активностью [87].
Недавняя научная литература идентифицирует IL-6 как важный регулятор опосредованного сателлитными клетками (мышечными стволовыми клетками) гипертрофического роста мышц. IL-6 локально и временно продуцируется растущими миофибриллами и ассоциированными сателлитными клетками, а генетическая потеря IL-6 притупляет мышечную гипертрофию in vivo [85]. Космиду и др. [86] показали, что АФК стимулируют высвобождение IL-6 из скелетных мышечных трубок, таким образом, идентифицируя новый клеточный источник, который может быть ответственным за избыточное количество цитокинов, чувствительных к воспалению, вырабатываемых в моделях с напряженными упражнениями.Их результаты также предполагают, что стимулированное АФК увеличение высвобождения IL-6 зависит от транскрипции и включает активацию p38 и NF𝜅-B. Хотя p38-MAPK появляется как ключевая молекула, управляющая последовательными событиями в миогенном пути, многие детали p38-MAPK-индуцированного миогенеза еще предстоит выяснить. Еще предстоит определить, регулируют ли различные члены семейства p38 MAPK экспрессию отдельных субнаборов генов на разных стадиях дифференцировки и обладают ли они индуцирующей или репрессирующей активностью [87].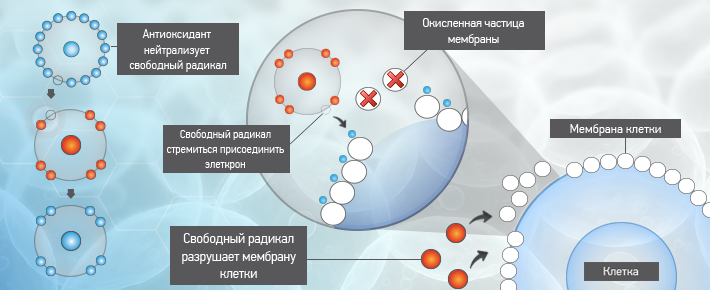
Баланс между производством реактивных частиц и системами антиоксидантной защиты, по-видимому, представляет собой центральный ключ многих адаптивных реакций в скелетных мышцах, а также гипертрофии и атрофии. Похоже, что если продукция АФК идеальна (чуть выше базового уровня), то адаптивной реакцией является гипертрофия; но если уровни АФК увеличиваются во много раз по сравнению с базальным уровнем и способностью к антиоксидантной защите, возникает атрофическая реакция, такая как мышечная дистрофия Дюшенна [88].
6. Выводы
Как было показано, сложная сигнально-транскрипционная сеть в мышечных волокнах опосредует вызванную физической нагрузкой адаптацию скелетных мышц, и имеются убедительные доказательства того, что АФК играют важную роль в этом сценарии. АФК участвуют в ангиогенезе, митохондриальном биогенезе и гипертрофии, которые играют важную роль в назначении упражнений и спортивных результатах. Что касается острых упражнений и образования свободных радикалов, как анаэробные, так и аэробные упражнения могут привести к увеличению образования свободных радикалов, но острый окислительный стресс не всегда может наблюдаться, поскольку производство АФК положительно зависит от нагрузки (интенсивность × продолжительность) упражнений. Было высказано предположение, что высокий уровень АФК, вызванный резкими приступами физических упражнений, может нанести ущерб иммунной системе; однако хронические упражнения вызывают физиологические адаптации, способные активизировать антиоксидантную систему человека. В связи с этим тип и продолжительность тренировки являются ключевыми для значительного повышения уровня эндогенных антиоксидантов, при этом длительные высокоинтенсивные тренировки на выносливость оказались более эффективными.
Было высказано предположение, что высокий уровень АФК, вызванный резкими приступами физических упражнений, может нанести ущерб иммунной системе; однако хронические упражнения вызывают физиологические адаптации, способные активизировать антиоксидантную систему человека. В связи с этим тип и продолжительность тренировки являются ключевыми для значительного повышения уровня эндогенных антиоксидантов, при этом длительные высокоинтенсивные тренировки на выносливость оказались более эффективными.
7. Перспективы на будущее
За последние 3 десятилетия мы значительно продвинулись в понимании влияния реактивных видов на адаптивную реакцию при физической нагрузке.Мышечные источники АФК были идентифицированы, и многие из сигнальных путей, которые модифицируются АФК, были изучены; однако многое еще предстоит открыть в отношении функции АФК в специфической адаптивной реакции, такой как ангиогенез, гипертрофия и митохондриальный биогенез. В настоящее время существует лучшее понимание регуляции источников АФК во время упражнений, но это все еще требует дальнейшего изучения. Необходима дополнительная работа для выяснения конкретной роли реактивных видов в процессах миграции, таких как перекатывание, прилипание и трансмиграция клеток; механистические процессы адаптации должны быть исследованы.Наконец, многие исследователи работают над расширением наших знаний о молекулярных и клеточных механизмах, связанных с взаимосвязью между окислительным стрессом и физическими упражнениями.
Необходима дополнительная работа для выяснения конкретной роли реактивных видов в процессах миграции, таких как перекатывание, прилипание и трансмиграция клеток; механистические процессы адаптации должны быть исследованы.Наконец, многие исследователи работают над расширением наших знаний о молекулярных и клеточных механизмах, связанных с взаимосвязью между окислительным стрессом и физическими упражнениями.
Химические, биологические и функциональные P
Содержание
Окисление продуктов питания: введение
Anna Kołakowska и Grzegorz Bartosz
Окислители, происходящие в пищевых системах
Manuel Pazos и Isabel Medina
Измерение потенциала окисления в продуктах питания
Луиза Беннетт, Эми Логан, Netsanet Shiferaw- Terefe, Tanoj Singh, and Robyn Warner
Механизмы окисления пищевых липидов
Gloria Márquez-Ruiz, Francisca Holgado, and Joaquín Velasco
Окисление белков в пищевых продуктах и его предотвращение 3 90 Caroline. Baron
Baron
Использование добавленных окислителей в пищевой промышленности
Emanuela Zanardi
Влияние окисления на сенсорные характеристики пищевых компонентов во время обработки и хранения
Susan Brewer
Пищевые компоненты
Rosario Zamora, Rosa M. Delgado, and Francisco J. Hidalgo
Натуральные антиоксиданты в пищевых системах
Petras Rimantas Venskutonis
Антиоксиданты, образующиеся в пищевых продуктах в результате обработки 3 3 Ibánez, и Miguel Herrero
Механизмы антиоксидантной активности
Klaudia Jomova, Michael Lawson и Marian Valko
Измерение антиоксидантной активности продуктов питания
Takayuki Shibamoto
Измерение антиоксидантной активности Apple Products
Iwona Вавер
Антиоксидантная и прокидантная активность пищевых компонентов
Anna Gliszczyńska-świgło и Ян Осмианьский
биодоступность и антиоксидантная активность куркуминоидов и каротиноидов в людях
Alexa Kocher, Кристина Шиброр, Даниэла Вебера, Тильман Грену и Ян Фрэнк
Тематические исследования отдельных натуральных пищевых антиоксидантов
Miguel Herrero, José A. Mendiola, Alejandro Cifuentes, и Elena Ibánez
Mendiola, Alejandro Cifuentes, и Elena Ibánez
функциональные антиоксидантные продукты
Manuel Viuda-Martos, Jose A. Pérez- Alvarez, и Juana Fernández-López
заключительные замечания
Grzegorz Bartosz и Izabela Sadowska-Bartosz
Значение антиоксидантов, играющих роль в клеточном ответе на окислительный/нитрозативный стресс: текущее состояние | Журнал о питании
Kurutas EB, Ciragil P, Gul M, Kilinc M.Последствия окислительного стресса при инфекциях мочевыводящих путей. Медиаторы воспаления. 2005; 2005: 242–4.
ПабМед Статья КАС Google Scholar
Валко М., Лейбфриц Д., Монкол Дж., Кронин М.Т., Мазур М., Телсер Дж. Свободные радикалы и антиоксиданты в нормальных физиологических функциях и заболеваниях человека. Int J Biochem Cell Biol. 2007; 39:44–84.
КАС пабмед Статья Google Scholar
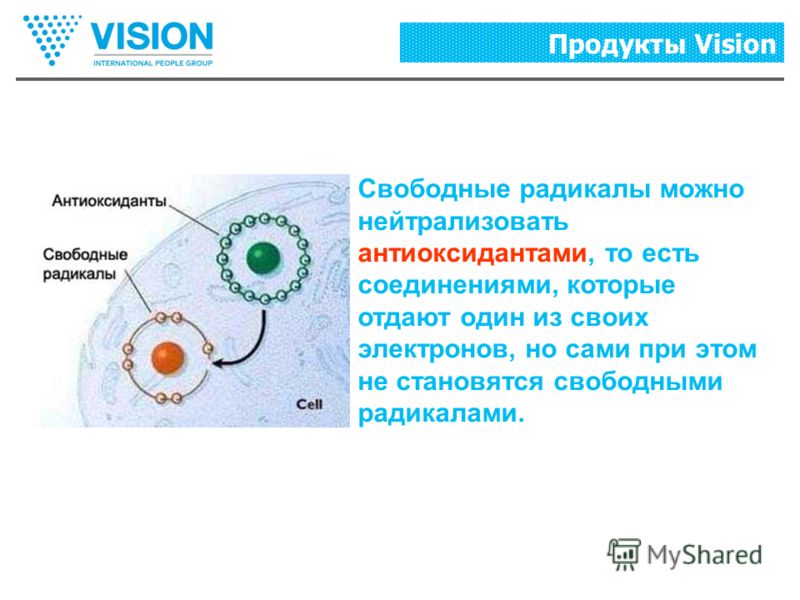
Барбаканн М.А., Сушард Дж.П., Дарблейд Б., Илиоу Дж.П., Непвеу Ф., Пипи Б., Баярд Ф., Арнал Дж.Ф. Обнаружение супероксид-аниона, высвобождаемого внеклеточно эндотелиальными клетками, с использованием методов восстановления цитохрома с, СОЭ, флуоресценции и люцигенин-усиленной хемилюминесценции. Свободный Радик Биол Мед. 2000; 29: 388–96.
КАС пабмед Статья Google Scholar
Vidrio E, Jung H, Anastasio C. Генерация гидроксильных радикалов из растворенных переходных металлов в суррогатных жидкостях легких.Атмос Окружающая среда. 2008; 42:4369–79.
КАС ПабМед Центральный Статья Google Scholar
Murphy E, Steenbergen C. Механизмы, лежащие в основе острой защиты от сердечной ишемии-реперфузии. Physiol Rev. 2008; 88: 581–609.
КАС пабмед ПабМед Центральный Статья Google Scholar
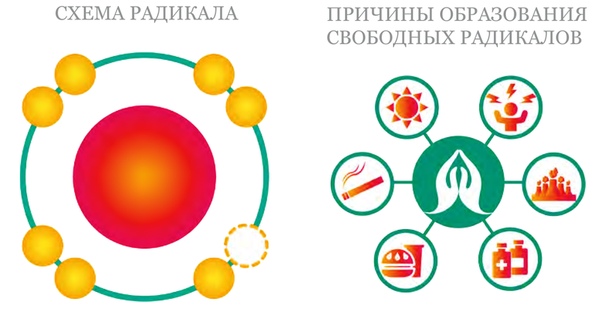
Бортолетто П., Лайман К., Камачо А., Фриккионе М., Ханолкар А., Кац Б.З.Хроническая гранулематозная болезнь. Pediatr Infect Dis J. 2015;34:1110–4.
ПабМед ПабМед Центральный Статья Google Scholar
Цао П.С., Хейдари С., Ван А., Чан Дж.Р., Ривен Г.М., Кук Д.П. Протеинкиназа С-эпсилон опосредует индуцированную глюкозой продукцию супероксида и экспрессию МСР-1 в эндотелиальных клетках. FASEB J. 1998; 12:512.
Google Scholar
Мастерс CJ.Клеточная передача сигналов: роль пероксисомы. Сотовый сигнал. 1996; 8: 197–208.
КАС пабмед Статья Google Scholar
Седербаум А.И. Молекулярные механизмы микросомальных оксидаз смешанной функции и биологические и патологические последствия. Редокс Биол. 2015;4:60–73.
КАС пабмед Статья Google Scholar

Dröge W. Свободные радикалы в физиологическом контроле клеточной функции.Physiol Rev. 2002; 82: 47–95.
ПабМед Статья Google Scholar
Валко М., Родс С.Дж., Монкол Дж., Изакович М.М., Мазура М. Свободные радикалы, металлы и антиоксиданты при раке, вызванном окислительным стрессом. Химическое биологическое взаимодействие. 2006; 160:1–40.
КАС пабмед Статья Google Scholar
Tatsuzawa H, Maruyama T, Hori K, Sano Y, Nakano M. Синглетный кислород ((1)Delta(g)O(2)) как основной окислитель в опосредованном миелопероксидазой уничтожении бактерий в фагосомах нейтрофилов.Biochim Biophys Res Commun. 1999; 262: 647–50.
КАС Статья Google Scholar
Ллойд Р.В., Ханна П.М., Мейсон Р.П. Происхождение гидроксильного радикала кислорода в реакции Фентона. Свободный Радик Биол Мед. 1997; 22:885–8.
1997; 22:885–8.
КАС пабмед Статья Google Scholar
Stohs SJ, Bagchi D. Окислительные механизмы при токсичности ионов металлов. Свободный Радик Биол Мед.1995; 18: 321–36.
КАС пабмед Статья Google Scholar
Пеппер Дж. Р., Мамби С., Гаттеридж Дж. К. Последовательное окислительное повреждение и изменения в железосвязывающих и железоокисляющих антиоксидантах плазмы во время операции искусственного кровообращения. Свободный Радик Рез. 1994; 21: 377–85.
КАС пабмед Статья Google Scholar
Джомова К., Валко М. Важность хелирования железа при окислительном стрессе, вызванном свободными радикалами, и заболеваниях человека.Курр Фарм Дез. 2011;17:3460–73.
КАС пабмед Статья Google Scholar

Saito H, Hayashi H. Скорость трансформации между ферритином и гемосидерином, определяемая кинетикой ферритина в сыворотке у пациентов с нормальными запасами железа и перегрузкой железом. Нагоя J Med Sci. 2015; 77: 571–83.
ПабМед ПабМед Центральный Google Scholar
Гафурифар П., Каденас Э.Митохондриальная синтаза оксида азота. Trends Pharmacol Sci. 2005; 26:190–5.
КАС пабмед Статья Google Scholar
Bergendi L, Benes L, Durackova Z, Ferencik M. Химия, физиология и патология свободных радикалов. Жизнь наук. 1999; 65: 1865–74.
КАС пабмед Статья Google Scholar
Чиуэ CC. Нейропротекторные свойства оксида азота.Энн NY Acad Sci. 1999; 890: 301–11.
КАС пабмед Статья Google Scholar

Beckman JS, Koppenol WH. Оксид азота, супероксид и пероксинитрит: хорошие, плохие и уродливые. Am J Physiol. 1996; 271:C1424–37.
КАС пабмед Google Scholar
Pacher P, Beckman JS, Liaudet L. Оксид азота и пероксинитрит в норме и при болезни.Physiol Rev. 2007; 87: 315–424.
КАС пабмед ПабМед Центральный Статья Google Scholar
Янг И.С., СП Вудсайд. Антиоксиданты в здоровье и болезни. Джей Клин Патол. 2001; 54: 176–86.
КАС пабмед ПабМед Центральный Статья Google Scholar
Флора SJ. Структурные, химические и биологические аспекты антиоксидантов для стратегий защиты от воздействия металлов и металлоидов.Оксид Мед Селл Лонгев. 2009;2:191–206.
ПабМед ПабМед Центральный Статья Google Scholar

Финкель Т., Холбрук, штат Нью-Джерси. Оксиданты, окислительный стресс и биология старения. Природа. 2000; 408: 239–47.
КАС пабмед Статья Google Scholar
Учида К. 4-гидрокси-2-ноненаль: продукт и медиатор окислительного стресса. Прог Липид Рез.2003;42:318–43.
КАС пабмед Статья Google Scholar
Карини М., Альдини Г., Фачино Р.М. Масс-спектрометрия для обнаружения аддуктов 4-гидрокси- транс--2-оненаля (HNE) с пептидами и белками. Масс-спектр, ред. 2004; 23: 281–305.
КАС пабмед Статья Google Scholar
Cracowski JL, Durand T, Bessard G. Изопростаны как биомаркеры перекисного окисления липидов у человека: физиология, фармакология и клиническое значение.Trends Pharmacol Sci. 2002; 23:360–6.
КАС пабмед Статья Google Scholar

Монтуши П., Барнс П.Дж., Робертс Л.Дж. Изопростаны: маркеры и медиаторы окислительного стресса. FASEB J. 2004; 18:1791–800.
КАС пабмед Статья Google Scholar
Учида К. Гистидин и лизин как мишени окислительной модификации. Аминокислоты.2003; 25: 249–57.
КАС пабмед Статья Google Scholar
Draper HH, Csallany AS, Hadley M. Альдегиды в моче как индикаторы перекисного окисления липидов in vivo. Свободный Радик Биол Мед. 2000;29:1071–7.
КАС пабмед Статья Google Scholar
Wilson R, Lyall K, Smyth L, Fernie CE, Riemersma RA. Пищевые гидроксижирные кислоты усваиваются людьми: последствия для измерения «окислительного стресса» in vivo.Свободный Радик Биол Мед. 2002; 32: 162–8.
КАС пабмед Статья Google Scholar

Басу С. Изопростаны: новые биоактивные продукты перекисного окисления липидов. Свободный Радик Рез. 2004; 38: 105–22.
КАС пабмед Статья Google Scholar
Miraloglu M, Kurutas EB, Ozturk P, Arican O. Оценка местного статуса микроэлементов и концентрации 8-изо-простагландина F2α у пациентов с Tinea pedis.Биол Проведено онлайн. 2016;18:1.
ПабМед ПабМед Центральный Статья Google Scholar
Морроу Д.Д. Количественное определение изопростанов как показателей оксидантного стресса и риска атеросклероза у людей. Артериосклеры Тромб Васк Биол. 2005; 25: 279–86.
КАС пабмед Статья Google Scholar
Морет-Татай И., Иборра М., Серрильо Э., Тортоса Л., Нос П., Бельтран Б.Возможные биомаркеры в крови для болезни Крона: окислительный стресс и микроРНК — текущие данные и дальнейшие аспекты, которые необходимо раскрыть. Оксид Мед Селл Лонгев. 2016;2016:2325162.
Оксид Мед Селл Лонгев. 2016;2016:2325162.
ПабМед Статья Google Scholar
Poli G, Leonarduzzi G, Biasi F, Chiarpotto E. Окислительный стресс и передача сигналов клетками. Курр Мед Хим. 2004; 11:1163–82.
КАС пабмед Статья Google Scholar
Eizirik DL, Manndrup-Poulsen T. Выбор смерти — сигнальная трансдукция иммуноопосредованного апоптоза бета-клеток. Диабетология. 2001;44:2115–33.
КАС пабмед Статья Google Scholar
Buttke TM, Sandstrom PA. Окислительный стресс как медиатор апоптоза. Иммунол сегодня. 1994; 15:1–4.
Артикул Google Scholar
Ники Э. Свободные радикалы в 1900-х годах: от in vitro к in vivo.Свободный Радик Рез. 2001; 33: 693–704.
Артикул Google Scholar

Клауниг Ю.Э., Камендулис Л.М., Хосевар Б.А. Окислительный стресс и окислительное повреждение при канцерогенезе. Токсикол патол. 2010; 38:96–109.
КАС пабмед Статья Google Scholar
Павлович Д., Кочич Р., Коцич Г., Евтович Т., Раденкович С., Микич Д., Стоянович М., Джорджевич П.Б. Влияние четырехнедельного лечения метформином на ферменты антиоксидантной защиты плазмы и эритроцитов у пациентов с недавно диагностированным ожирением NIDDM.Сахарный диабет Ожирение Metab. 2000; 2: 251–6.
КАС пабмед Статья Google Scholar
Диас М.Н., Фрей Б., Вита Дж.А., Кини Дж.Ф. Антиоксиданты и атеросклеротическая болезнь сердца. N Engl J Med. 1997; 337: 408–16.
КАС пабмед Статья Google Scholar
Marinho HS, Real C, Cyrne L, Soares H, Antunes F. Регулирование факторов восприятия, передачи сигналов и транскрипции перекиси водорода.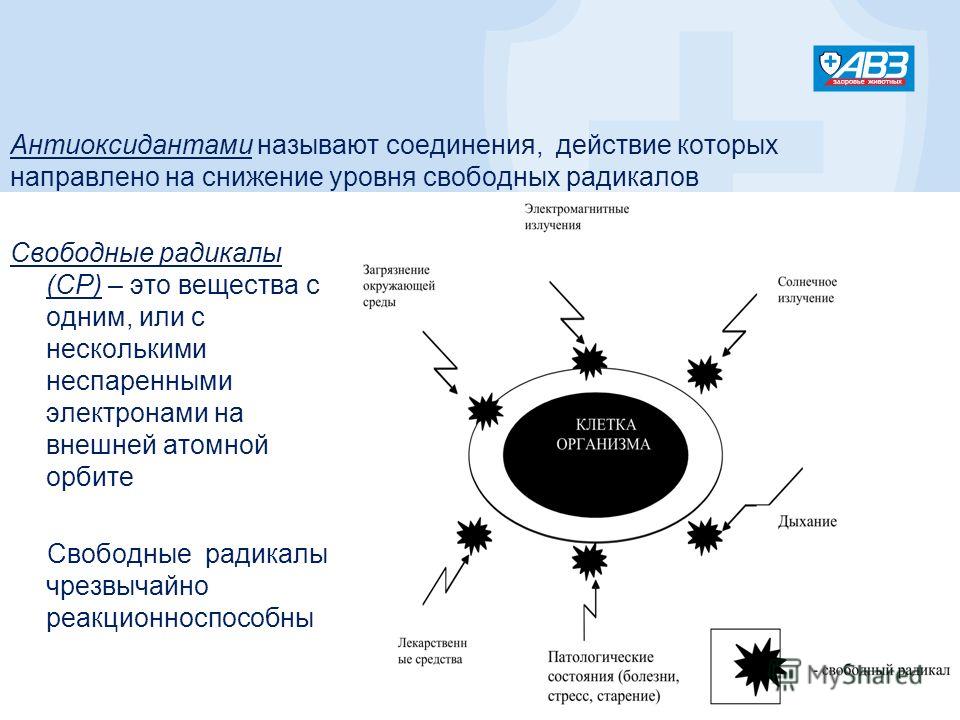 Редокс Биол. 2014;2:535–62.
Редокс Биол. 2014;2:535–62.
КАС пабмед ПабМед Центральный Статья Google Scholar
Nabavi SF, Barber AJ, Spagnuolo C, Russo GL, Daglia M, Nabavi SM, Sobarzo-Sánchez E. Nrf2 как молекулярная мишень для полифенолов: новая терапевтическая стратегия при диабетической ретинопатии. Crit Rev Clin Lab Sci. 2016; 29:1–20.
Google Scholar
Пан С, Шэн И, Цзян П, Вэй Х, Цзи Л.Хлорогеновая кислота предотвращает повреждение печени, вызванное ацетаминофеном: участие метаболических ферментов CYP450 и некоторых антиоксидантных сигналов. J Zhejiang Univ Sci B. 2015; 16: 602–10.
КАС пабмед ПабМед Центральный Статья Google Scholar
Прасад КН. Одновременная активация Nrf2 и повышение уровня пищевых и эндогенных антиоксидантных химических веществ для профилактики рака у людей.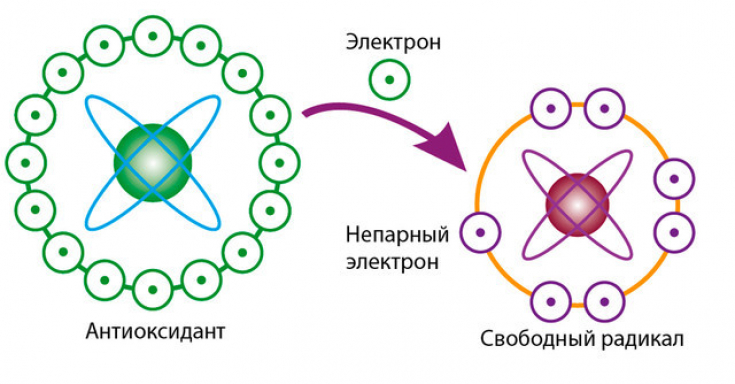 J Am Coll Nutr. 2016; 35:175–84.
J Am Coll Nutr. 2016; 35:175–84.
КАС пабмед Статья Google Scholar
Чан К., Хан XD, Кан Ю.В. Важная функция Nrf2 в борьбе с окислительным стрессом: детоксикация ацетаминофена. Proc Natl Acad Sci U S A. 2001;98:4611–6.
КАС пабмед ПабМед Центральный Статья Google Scholar
Enomoto A, Itoh K, Nagayoshi E, Haruta J, Kimura T, O’Connor T, Harada T, Yamamoto M. Высокая чувствительность мышей с нокаутом Nrf2 к гепатотоксичности ацетаминофена, связанная со снижением экспрессии ARE-регулируемого препарата метаболизирующие ферменты и антиоксидантные гены.Токсикол науч. 2001; 59: 169–77.
КАС пабмед Статья Google Scholar
Чан К., Кан Ю.В. Nrf2 необходим для защиты от острого повреждения легких у мышей. Proc Natl Acad Sci U S A. 1999;96:12731–6.
КАС пабмед ПабМед Центральный Статья Google Scholar

Fahey JW, Haristoy X, Dolan PM, Kensler TW, Scholtus I, Stephenson KK, Talalay P, Lozniewski A.Сульфорафан ингибирует внеклеточные, внутриклеточные и устойчивые к антибиотикам штаммы Helicobacter pylori и предотвращает опухоли желудка, вызванные бензо[а]пиреном. Proc Natl Acad Sci U S A. 2002;99:7610–5.
КАС пабмед ПабМед Центральный Статья Google Scholar
Рамос-Гомез М., Квак М.К., Долан П.М., Ито К., Ямамото М., Талалай П., Кенслер Т.В. У мышей с дефицитом транскрипционного фактора nrf2 повышена чувствительность к канцерогенезу и утрачена химиозащитная эффективность индукторов ферментов.Proc Natl Acad Sci U S A. 2001;98:3410–5.
КАС пабмед ПабМед Центральный Статья Google Scholar
Braun S, Hanselmann C, Gassmann MG, Auf Dem Keller U, Born-Berclaz C, Chan K, Kan YW, Werner S. Фактор транскрипции Nrf2, новая мишень действия фактора роста кератиноцитов, который регулирует экспрессию генов и воспаление в заживающей кожной ране. Мол Селл Биол. 2002; 22: 5492–505.
Мол Селл Биол. 2002; 22: 5492–505.
КАС пабмед ПабМед Центральный Статья Google Scholar
Дакшинамурти С., Лонг Д.Дж., Джайсвал А.К. Антиоксидантная регуляция генов, кодирующих ферменты, обезвреживающие ксенобиотики и канцерогены. Curr Top Cell Regul. 2000; 36: 201–16.
КАС пабмед Статья Google Scholar
Ли Дж.М., Меленкамп Дж.Д., Хэнсон Дж.М., Джонсон Дж.А. Nrf2-зависимая активация антиоксидантного чувствительного элемента трет-бутилгидрохиноном не зависит от окислительного стресса в клетках нейробластомы человека IMR-32.Biochem Biophys Res Commun. 2001; 280: 286–92.
КАС пабмед Статья Google Scholar
Ли Дж.М., Хэнсон Дж.М., Чу В.А., Джонсон Дж.А. Фосфатидилинозитол-3-киназа, а не киназа, регулируемая внеклеточным сигналом, регулирует активацию антиоксидант-чувствительного элемента в клетках нейробластомы человека IMR-32. Дж. Биол. Хим. 2001; 276:20011–6.
Дж. Биол. Хим. 2001; 276:20011–6.
КАС пабмед Статья Google Scholar
Alam J, Wicks C, Stewart D, Gong P, Touchard C, Otterbein S, Choi AM, Burow ME, Tou J. Механизм активации гена гемоксигеназы-1 кадмием в эпителиальных клетках молочной железы MCF-7. Роль киназы p38 и фактора транскрипции Nrf2. Дж. Биол. Хим. 2000; 275:27694–702.
КАС пабмед Google Scholar
Kim YC, Masutani H, Yamaguchi Y, Itoh K, Yamamoto M, Yodoi J. Индуцированная гемином активация гена тиоредоксина с помощью Nrf2.Дифференциальная регуляция антиоксидантного чувствительного элемента путем переключения факторов его связывания. Дж. Биол. Хим. 2001; 276:18399–406.
КАС пабмед Статья Google Scholar
Орино К., Леман Л., Цудзи Ю., Аяки Х., Торти С.В., Торти FM. Ферритин и реакция на окислительный стресс. Биохим Дж. 2001;357:241–7.
Биохим Дж. 2001;357:241–7.
КАС пабмед ПабМед Центральный Статья Google Scholar
Motohashi H, O’Connor T, Katsuoka F, Engel JD, Yamamoto M. Интеграция и разнообразие регуляторной сети, состоящей из семейств транскрипционных факторов Maf и CNC. Ген. 2002; 294:1–12.
КАС пабмед Статья Google Scholar
Chanas SA, Jiang Q, McMahon M, McWalter GK, McLellan LI, Elcombe CR, Henderson CJ, Wolf CR, Moffat GJ, Itoh K, Yamamoto M, Hayes JD. Потеря фактора транскрипции Nrf2 вызывает заметное снижение конститутивной и индуцибельной экспрессии генов глутатион-S-трансферазы Gsta1, Gsta2, Gstm1, Gstm2, Gstm3 и Gstm4 в печени самцов и самок мышей.Биохим Дж. 2002;365:405–16.
КАС пабмед ПабМед Центральный Статья Google Scholar
Hayes JD, Chanas SA, Henderson CJ, McMahon M, Sun C, Moffat GJ, Wolf CR, Yamamoto M. Фактор транскрипции Nrf2 способствует как базовой экспрессии глутатион-S-трансфераз в печени мыши, так и их индукция химиопрофилактическими синтетическими антиоксидантами, бутилированным гидроксианизолом и этоксихином. Биохим Сок Транс.2000; 28:33–41.
Фактор транскрипции Nrf2 способствует как базовой экспрессии глутатион-S-трансфераз в печени мыши, так и их индукция химиопрофилактическими синтетическими антиоксидантами, бутилированным гидроксианизолом и этоксихином. Биохим Сок Транс.2000; 28:33–41.
КАС пабмед Статья Google Scholar
Ishii T, Itoh K, Takahashi S, Sato H, Yanagawa T, Katoh Y, Bannai S, Yamamoto M. Фактор транскрипции Nrf2 координированно регулирует группу генов, индуцируемых окислительным стрессом, в макрофагах. Дж. Биол. Хим. 2000; 275:16023–9.
КАС пабмед Статья Google Scholar
Нгуен Т., Хуанг Х.К., Пикетт С.Б.Транскрипционная регуляция элемента антиоксидантной реакции. Активация Nrf2 и подавление MafK. Дж. Биол. Хим. 2000; 275:15466–73.
КАС пабмед Статья Google Scholar
Wild AC, Moinova HR, Mulcahy RT. Регуляция экспрессии гена субъединицы гамма-глутамилцистеинсинтетазы фактором транскрипции Nrf2. Дж. Биол. Хим. 1999; 274:33627–36.
Регуляция экспрессии гена субъединицы гамма-глутамилцистеинсинтетазы фактором транскрипции Nrf2. Дж. Биол. Хим. 1999; 274:33627–36.
КАС пабмед Статья Google Scholar
Алам Дж., Стюарт Д., Тушар С., Бойнапалли С., Чой А.М., Кук Дж.Л. Nrf2, транскрипционный фактор Cap’n’Collar, регулирует индукцию гена гемоксигеназы-1. Дж. Биол. Хим. 1999; 274:26071–8.
КАС пабмед Статья Google Scholar
Yu R, Lei W, Mandlekar S, Weber MJ, Der CJ, Wu J, Kong AN. Роль митоген-активируемого протеинкиназного пути в индукции ферментов детоксикации фазы II химическими веществами. Дж. Биол. Хим.1999; 274:27545–52.
КАС пабмед Статья Google Scholar
Huang HC, Nguyen T, Pickett CB. Регуляция элемента антиоксидантной реакции с помощью опосредованного протеинкиназой C фосфорилирования фактора 2, родственного NF-E2. Proc Natl Acad Sci U S A. 2000; 97:12475–8031.
Proc Natl Acad Sci U S A. 2000; 97:12475–8031.
КАС пабмед ПабМед Центральный Статья Google Scholar
Ли Дж., Ли Дж.М., Джонсон Дж.А.Анализ микрочипов выявил набор генов, управляемых элементами, чувствительными к антиоксидантам, которые участвуют в обеспечении защиты от апоптоза, вызванного окислительным стрессом, в клетках IMR-32. Дж. Биол. Хим. 2002; 277: 388–94.
КАС пабмед Статья Google Scholar
Ли Дж., Джонсон Дж.А. Зависимые от времени изменения в экспрессии генов, управляемой ARE, с использованием процесса фильтрации шума для данных микрочипов. Физиол Геномика. 2002; 9: 137–44.
ПабМед Статья Google Scholar
Джонсон Д.А., Эндрюс Г.К., Сюй В., Джонсон Дж.А. Активация элемента антиоксидантной реакции в первичных культурах нейронов коры головного мозга, полученных от трансгенных репортерных мышей. Дж. Нейрохим. 2002; 81: 1233–41.
Дж. Нейрохим. 2002; 81: 1233–41.
КАС пабмед Статья Google Scholar
Кан К.В., Чо М.К., Ли Ч., Ким С.Г. Активация фосфатидилинозитол-3-киназы и Akt трет-бутилгидрохиноном отвечает за опосредованную антиоксидантным элементом индукцию rGSTA2 в клетках h5IIE.Мол Фармакол. 2001; 59: 1147–56.
КАС пабмед Google Scholar
Ли Дж.М., Джонсон Дж.А. Важная роль пути Nrf2-ARE в механизме клеточной защиты. Дж. Биохим Мол. Биол. 2004; 37: 139–43.
КАС пабмед Google Scholar
Suzuki T, Yamamoto M. Молекулярная основа системы Keap1-Nrf2. Свободный Радик Биол Мед. 2015; 88: 93–100.
КАС пабмед Статья Google Scholar
Нитуре СК, Хатри Р., Джайсвал АК. Регламент Nrf2 – обновление. Свободный Радик Биол Мед. 2014;66:34–6.
2014;66:34–6.
Артикул КАС Google Scholar
Форман Х.Дж., Дэвис К.Дж.А., Урсини Ф. Как на самом деле работают пищевые антиоксиданты: нуклеофильный тонус и парагормезис, удаляющий свободные радикалы in vivo. Свободный Радик Биол Мед. 2014;66:24–35.
КАС пабмед Статья Google Scholar
Внуков В.В., Гуценко О.И., Милютина Н.П., Ананян А.А., Даниленко А.О., Панина С.Б., Корниенко И.В. Влияние SkQ1 на экспрессию гена фактора транскрипции Nrf2, ARE-контролируемых генов антиоксидантных ферментов и их активность в лейкоцитах крови крыс. Биохимия (Москва). 2015; 80: 586–91.
КАС Статья Google Scholar
Ито К., Мимура Дж., Ямамото М. Открытие отрицательного регулятора Nrf2, Keap1: исторический обзор.Антиоксидный окислительно-восстановительный сигнал. 2010; 13:1665–78.
КАС пабмед Статья Google Scholar

Kensler TW, Wakabayashi N. Nrf2: друг или враг для химиопрофилактики? Канцерогенез. 2010;31:90–9.
КАС пабмед Статья Google Scholar
Солис Л.М., Беренс С., Донг В., Сураокар М., Озберн Н.К., Моран К.А., Корвалан А.Х., Бисвал С., Свишер С.Г., Бекеле Б.Н., Минна Д.Д., Стюарт Д.Дж., Вистуба II.Аномалии Nrf2 и Keap1 при немелкоклеточной карциноме легкого и связь с клинико-патологическими особенностями. Клин Рак Рез. 2010;16:3743–53.
КАС пабмед ПабМед Центральный Статья Google Scholar
Сингх А., Мисра В., Тиммулаппа Р.К., Ли Х., Эймс С., Хок М.О., Герман Дж.Г., Байлин С.Б., Сидрански Д., Габриэльсон Э., Брок М.В., Бисвал С. Дисфункциональное взаимодействие KEAP1-NRF2 в не- мелкоклеточный рак легкого. ПЛОС Мед.2006;3:e420.
ПабМед ПабМед Центральный Статья КАС Google Scholar

Shibata T, Kokubu A, Gotoh M, Ojima H, Ohta T, Yamamoto M, Hirohashi S. Генетическое изменение Keap1 обеспечивает конститутивную активацию Nrf2 и устойчивость к химиотерапии при раке желчного пузыря. Гастроэнтерол. 2008; 135:1358–68. 1368.e1–4.
КАС Статья Google Scholar
Li K, Zhong C, Wang B, He J, Bi J. Экспрессия Nrf2 участвует в росте и дифференцировке клеток карциномы эндометрия in vitro и in vivo. Дж. Мол. Хистол. 2014;45:161–7.
ПабМед Статья КАС Google Scholar
Zhang P, Singh A, Yegnasubramanian S, Esopi D, Kombairaju P, Bodas M, Wu H, Bova SG, Biswal S. Потеря функции Kelch-подобного ECH-ассоциированного белка 1 в клетках рака предстательной железы вызывает химиорезистентность и радиорезистентность и способствует росту опухоли.Мол Рак Тер. 2010;9:336–46.
КАС пабмед ПабМед Центральный Статья Google Scholar

Ji XJ, Chen SH, Zhu L, Pan H, Zhou Y, Li W, You WC, Gao CC, Zhu JH, Jiang K, Wang HD. Нокдаун фактора 2, родственного NF-E2, ингибирует пролиферацию и рост клеток глиомы человека U251MG в модели ксенотрансплантата мыши. Oncol Rep. 2013; 30:157–64.
КАС пабмед Google Scholar
Soini Y, Eskelinen M, Juvonen P, Kärjä V, Haapasaari KM, Saarela A, Karihtala P. Ядерная экспрессия Nrf2 связана с плохой выживаемостью при аденокарциноме поджелудочной железы. Патол Res Pract. 2014; 210:35–9.
КАС пабмед Статья Google Scholar
Кансанен Э., Куосманен С.М., Лейнонен Х., Левонен А.Л. Путь Keap1-Nrf2: механизмы активации и нарушения регуляции при раке. Редокс Биол. 2013;1:45–9.
КАС пабмед ПабМед Центральный Статья Google Scholar
Mitsuishi Y, Taguchi K, Kawatani Y, Shibata T, Nukiwa T, Aburatani H, Yamamoto M, Motohashi H. Nrf2 перенаправляет глюкозу и глютамин в анаболические пути при метаболическом перепрограммировании. Раковая клетка. 2012;22:66–79.
Nrf2 перенаправляет глюкозу и глютамин в анаболические пути при метаболическом перепрограммировании. Раковая клетка. 2012;22:66–79.
КАС пабмед Статья Google Scholar
Lennicke C, Rahn J, Lichtenfels R, Wessjohann LA, Seliger B. Редокс-протеомика: методы идентификации и обогащения редокс-модифицированных белков и их применение.Сигнал сотовой связи. 2015;13:39.
ПабМед ПабМед Центральный Статья КАС Google Scholar
Li L, Fath MA, Scarbrough PM, Watson WH, Spitz DR. Комбинированное ингибирование гликолиза, пентозного цикла и метаболизма тиоредоксина избирательно увеличивает цитотоксичность и окислительный стресс при раке молочной железы и предстательной железы человека. Редокс Биол. 2014; 4С: 127–35.
Google Scholar
Ji X, Wang H, Zhu J, Zhu L, Pan H, Li W, Zhou Y, Cong Z, Yan F, Chen S. Нокдаун Nrf2 подавляет ангиогенез глиобластомы путем ингибирования индуцированной гипоксией активации HIF-1α. Инт Джей Рак. 2014; 135:574–84.
Нокдаун Nrf2 подавляет ангиогенез глиобластомы путем ингибирования индуцированной гипоксией активации HIF-1α. Инт Джей Рак. 2014; 135:574–84.
КАС пабмед Статья Google Scholar
Карки Р., Игве О.Дж. Опосредованная Toll-подобным рецептором 4 активация ядерного фактора каппа B необходима для восприятия экзогенных оксидантов для распространения и поддержания окислительного/нитрозативного клеточного стресса.ПЛОС Один. 2013;8:e73840.
КАС пабмед ПабМед Центральный Статья Google Scholar
Gasparini C, Celeghini C, Monasta L, Zauli G. Пути NF-kB при гематологических злокачественных новообразованиях. Cell Mol Life Sci. 2014;71:2083–102.
КАС пабмед Статья Google Scholar
Ямамото Ю., Гейнор Р.Б. Роль пути NF-kB в патогенезе болезненных состояний человека. Курр Мол Мед. 2001; 1: 287–96.
Курр Мол Мед. 2001; 1: 287–96.
КАС пабмед Статья Google Scholar
Гош С., Мэй М.Дж., Копп Э.Б. Белки NF-каппа B и Rel: эволюционно консервативные медиаторы иммунных ответов. Анну Рев Иммунол. 1998; 16: 225–60.
КАС пабмед Статья Google Scholar
Oliveira-Marques V, Marinho HS, Cyrne L, Antunes F. Роль перекиси водорода в активации NF-kappaB: от индуктора к модулятору.Антиоксидный окислительно-восстановительный сигнал. 2009;11:2223–43.
КАС пабмед Статья Google Scholar
Jung Y, Kim H, Min SH, Rhee SG, Jeong W. Легкая цепь LC8 динеина отрицательно регулирует NF-kappaB посредством редокс-зависимого взаимодействия с IkappaBalpha. Дж. Биол. Хим. 2008; 283:23863–71.
КАС пабмед ПабМед Центральный Статья Google Scholar

Lee DF, Kuo HP, Liu M, Chou CK, Xia W, Du Y, Shen J, Chen CT, Huo L, Hsu MC, Li CW, Ding Q, Liao TL, Lai CC, Lin AC, Chang YH, Tsai С.Ф., Ли Л.И., Хунг М.С. Опосредованное лигазой KEAP1 E3 подавление передачи сигналов NF-kappaB путем нацеливания на IKKbeta. Мол Ячейка. 2009; 36: 131–40.
КАС пабмед ПабМед Центральный Статья Google Scholar
Фам К.Г., Бубичи С., Заззерони Ф., Папа С., Джонс Дж., Альварес К., Джаявардена С., Де Смаэле Э., Конг Р., Бомонт С., Торти FM, Торти С.В., Францозо Г.Активация тяжелой цепи ферритина с помощью NF-kappaB ингибирует апоптоз, индуцированный TNF-альфа, путем подавления активных форм кислорода. Клетка. 2004; 119: 529–42.
КАС пабмед Статья Google Scholar
Камата Х., Хонда С., Маэда С., Чанг Л., Хирата Х., Карин М. Активные формы кислорода способствуют гибели, вызванной альфа-ФНО, и устойчивой активации JNK путем ингибирования MAP-киназных фосфатаз. Клетка. 2005; 120:649–61.
Клетка. 2005; 120:649–61.
КАС пабмед Статья Google Scholar
Rius J, Guma M, Schachtrup C, Akassoglou K, Zinkernagel AS, Nizet V, Johnson RS, Haddad GG, Karin M. NF-kappaB связывает врожденный иммунитет с гипоксической реакцией посредством транскрипционной регуляции HIF-1alpha. Природа. 2008; 453:807–11.
КАС пабмед ПабМед Центральный Статья Google Scholar
Ma Q, Kinneer K, Ye J, Chen BJ. Ингибирование ядерного фактора kappaB фенольными антиоксидантами: взаимодействие между антиоксидантной сигнализацией и экспрессией воспалительных цитокинов.Мол Фармакол. 2003; 64: 211–9.
КАС пабмед Статья Google Scholar
Feng X, Tan W, Cheng S, Wang H, Ye S, Yu C, He Y, Zeng J, Cen J, Hu J, Zheng R, Zhou Y. Активация микроРНК-126 в звездчатой печенке клетки могут влиять на патогенез фиброза печени через путь NF-B. ДНК-клеточная биол. 2015; 34: 470–80.
КАС пабмед Статья Google Scholar
Коллинз Т., Цибульский М.И. NF-kB: ключевой медиатор или невинный наблюдатель атерогенеза? Джей Клин Инвест. 2001; 107: 255–64.
КАС пабмед ПабМед Центральный Статья Google Scholar
Ghashghaeinia M, Toulany M, Saki M, Rodemann HP, Mrowietz U, Lang F, Wieder T. Потенциальные роли путей NFkB и глутатиона в зрелых эритроцитах человека. Cell Мол Биол Летт. 2012; 17:11–20.
КАС Статья Google Scholar
Никола Дж.П., Пейре В., Назар М., Ромеро Дж.М., Лусеро А.М., Монтесинос Мдель М., Бокко Дж.Л., Пеллизас К.Г. Masini-Repiso AM S-нитрозилирование NF-kB p65 ингибирует индуцированную ТТГ экспрессию Na(+)/I(-) симпортера. Эндокринология. 2015; 156:4741–54.
КАС пабмед Статья Google Scholar

Shi X, Dong Z, Huang C, Ma W, Liu K, Ye J, Chen F, Leonard SS, Ding M, Castranova V, Vallyathan V. Роль гидроксильного радикала в качестве мессенджера в активации ядерного фактора транскрипции NF-kappaB.Мол Селл Биохим. 1999; 194: 63–70.
КАС пабмед Статья Google Scholar
Rahman I, Marwick J, Kirkham P. Окислительно-восстановительная модуляция ремоделирования хроматина: влияние на ацетилирование и деацетилирование гистонов, NF-kappaB и экспрессию провоспалительных генов. Биохим Фармакол. 2004; 68: 1255–67.
КАС пабмед Статья Google Scholar
Ямамото Ю., Гейнор Р.Б.Терапевтический потенциал ингибирования пути NF-kB при лечении воспаления и рака. Джей Клин Инвест. 2001; 107: 135–42.
КАС пабмед ПабМед Центральный Статья Google Scholar
Karin M, Takahashi T, Kapahi P, Delhase M, Chen Y, Makris C, Rothwarf D, Baud V, Natoli G, Guido F, Li N. Окислительный стресс и экспрессия генов: AP-1 и соединения NF-kB. Биофакторы. 2001; 15:87–89.
Окислительный стресс и экспрессия генов: AP-1 и соединения NF-kB. Биофакторы. 2001; 15:87–89.
КАС пабмед Статья Google Scholar
Уитмарш А.Дж., Дэвис Р.Дж. Регуляция фактора транскрипции AP-1 с помощью путей передачи сигнала митоген-активируемой протеинкиназы. J Mol Med (Берл). 1996;74(10):589–607.
КАС Статья Google Scholar
Пинкус Р., Вайнер Л.М., Даниэль В. Роль оксидантов и антиоксидантов в индукции экспрессии генов AP-1, NF-kB и глутатион-S-трансферазы. Дж. Биол. Хим. 1996; 271:13422–9.
КАС пабмед Статья Google Scholar
Павлович Д., Джорджевич В., Коцич Г. «Взаимодействие» между окислительным стрессом и передачей сигналов окислительно-восстановительных клеток. Серия Facta Unıv: Med Biol. 2002; 9: 131–7.
Google Scholar

Go YM, Patel RP, Maland MC, Park H, Beckman JS, Darley-Usmar VM, Jo H. Доказательства пероксинитрита как сигнальной молекулы в зависимой от потока активации c-Jun NH(2)- терминальная киназа. Am J Physiol. 1999; 277:h2647–1653.
КАС пабмед Google Scholar
Lander HM, Ogiste JS, Teng KK, Novogrodsky A. p21 ras как общая сигнальная мишень реактивных свободных радикалов и клеточного окислительно-восстановительного стресса. Дж. Биол. Хим. 1995; 270:21195–8.
КАС пабмед Статья Google Scholar
Накамура Х., Накамура К., Йодои Дж. Окислительно-восстановительная регуляция клеточной активации. Энн Рев Иммунол. 1997; 15: 351–69.
КАС Статья Google Scholar
Circu ML, Aw TY. Активные формы кислорода, клеточные окислительно-восстановительные системы и апоптоз. Свободный Радик Биол Мед. 2010;48:749–62.
КАС пабмед ПабМед Центральный Статья Google Scholar
Trachootham D, Lu W, Ogasawara MA, Valle NR, Huang P. Окислительно-восстановительная регуляция выживания клеток. Антиоксидный окислительно-восстановительный сигнал. 2008; 10:1343–74.
КАС пабмед ПабМед Центральный Статья Google Scholar
Коскун М., Бьеррум Дж.Т., Сейделин Дж.Б., Нильсен О.Х. МикроРНК при воспалительных заболеваниях кишечника — патогенез, диагностика и терапия. Мир J Гастроэнтерол. 2012;18:4629–34.
КАС пабмед ПабМед Центральный Статья Google Scholar
Leung AKL, Sharp PA. Функции микроРНК в реакциях на стресс. Мол Ячейка. 2010;40:205–15.
КАС пабмед ПабМед Центральный Статья Google Scholar

Lin Y, Liu X, Cheng Y, Yang J, Huo Y, Zhang C. Участие микроРНК в генной регуляции, опосредованной перекисью водорода, и ответе на клеточное повреждение в гладкомышечных клетках сосудов. Дж. Биол. Хим. 2009; 284:7903–13.
КАС пабмед ПабМед Центральный Статья Google Scholar
Wang L, Huang H, Fan Y, Kong B, Hu H, Hu K, Guo J, Mei Y и Liu W. Влияние подавления микроРНК-181a на h3O2-индуцированный апоптоз клеток H9c2 посредством митохондриальный путь апоптоза.Оксид Мед Селл Лонгев. 2014;2014:1–16.
Google Scholar
Christian P, Su Q. Регуляция микроРНК митохондриальных и ER сигнальных путей стресса: последствия для метаболизма липопротеинов при метаболическом синдроме. Am J Physiol—Endocrinol Metab. 2014; 307:E729–37.
КАС пабмед Статья Google Scholar
Маджента А.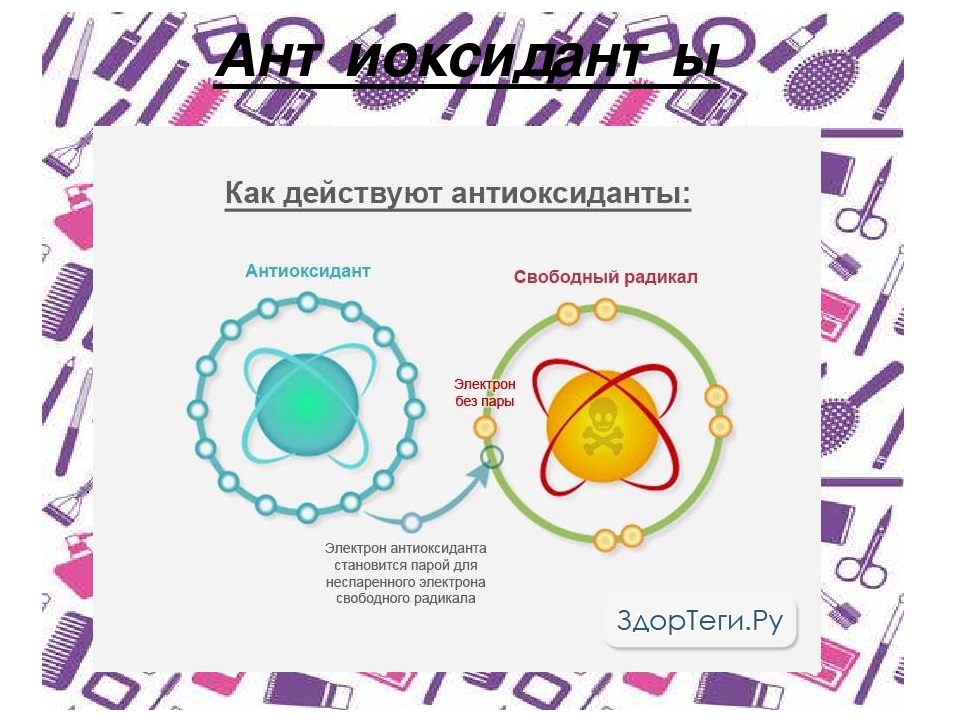 , Греко С., Гаэтано С., Мартелли Ф.Окислительный стресс и микроРНК при сосудистых заболеваниях. Int J Mol Sci. 2013; 14:17319–46.
, Греко С., Гаэтано С., Мартелли Ф.Окислительный стресс и микроРНК при сосудистых заболеваниях. Int J Mol Sci. 2013; 14:17319–46.
ПабМед ПабМед Центральный Статья КАС Google Scholar
Sies H. Окислительный стресс: концепция окислительно-восстановительной биологии и медицины. Редокс Биол. 2015;4:180–3.
КАС пабмед ПабМед Центральный Статья Google Scholar
Лопес-Бернардо Э., Анедда А., Санчес-Перес П., Акоста-Иборра Б., Каденас С.4-гидроксиноненаль индуцирует Nrf2-опосредованную активацию UCP3 в кардиомиоцитах мыши. Свободный Радик Биол Мед. 2015; 88: 427–38.
КАС пабмед Статья Google Scholar
Джонс Д.П., Сиес Х. Редокс-код. Антиоксидный окислительно-восстановительный сигнал. 2015; 23:734–46.
КАС пабмед ПабМед Центральный Статья Google Scholar

den Hertog J, Groen A, van der Wijk T.Окислительно-восстановительная регуляция протеинтирозинфосфатаз. Арх Биохим Биофиз. 2005; 434:11–5.
Артикул КАС Google Scholar
Gopalakrishna R, Jaken S. Передача сигналов протеинкиназы C и окислительный стресс. Свободный Радик Биол Мед. 2000; 28:1349–61.
КАС пабмед Статья Google Scholar
Zarubin T, Han J. Активация и сигнализация пути киназы p38 MAP.Сотовый рез. 2005; 15:11–8.
КАС пабмед Статья Google Scholar
Катарци С., Романьоли С., Маркуччи Г., Фавилли Ф., Янтомаси Т., Винчензини МТ. Окислительно-восстановительная регуляция активации ERK1/2, индуцированной сфингозин-1-фосфатом в фибробластах: участие НАДФН-оксидазы и рецептора тромбоцитарного фактора роста. Биохим Биофиз Акта. 1810; 2011: 446–56.
Google Scholar

Meng D, Shi X, Jiang BH, Fang J. Инсулиноподобный фактор роста-I (IGF-I) индуцирует трансактивацию рецептора эпидермального фактора роста и пролиферацию клеток посредством активных форм кислорода. Свободный Радик Биол Мед. 2007; 42:1651–60.
КАС пабмед Статья Google Scholar
Nagai H, Noguchi T, Takeda K, Ichijo H. Патофизиологические роли сигнальных путей киназы ASK1-MAP. Дж. Биохим Мол. Биол. 2007; 40:1–6.
КАС пабмед Google Scholar
Son Y, Cheong Y, Kim N, Chung H, Kang DG, Pae H. Митоген-активированные протеинкиназы и активные формы кислорода: как АФК могут активировать пути MAPK? J Сигнальный преобразователь. 2011;2011:1–6.
Артикул КАС Google Scholar
Чжоу ДЖИ, Лю Ю, Ву ГС. Роль митоген-активируемой протеинкиназы фосфатазы-1 в гибели клеток, вызванной окислительным повреждением. Рак рез. 2006; 66: 4888–94.
Рак рез. 2006; 66: 4888–94.
КАС пабмед Статья Google Scholar
Нитианандараджа-Джонс Г.Н., Вилм Б., Goldring CEP, Мюллер Дж., Кросс М.Дж. ERK5: Структура, регулирование и функция. Сотовый сигнал. 2012;24:2187–96.
КАС пабмед Статья Google Scholar
Бервелл Л.С., Надточий С.М., Томпкинс А.Дж., Янг С., Брукс П.С. Прямые доказательства S-нитрозирования митохондриального комплекса I. Biochem J. 2006;394:627–34.
КАС пабмед ПабМед Центральный Статья Google Scholar
Колесник Б., Палтен К., Шраммель А., Стессель Х., Шмидт К., Майер Б., Горрен АКФ. Эффективное нитрозирование глутатиона оксидом азота. Свободный Радик Биол Мед. 2013;63:51–64.
КАС пабмед ПабМед Центральный Статья Google Scholar

Эспиноса-Диес С., Мигель В., Меннерих Д., Китцманн Т., Санчес-Перес П., Каденас С., Ламас С. Антиоксидантные реакции и адаптация клеток к окислительному стрессу. Редокс Биол. 2015;6:183–97.
КАС пабмед ПабМед Центральный Статья Google Scholar
Абелло Н., Керстенс Х.А., Постма Д.С., Бишофф Р. Нитрование тирозина белков: селективность, физико-химические и биологические последствия, денитрование и протеомные методы идентификации белков, нитрованных тирозином. J Протеом Res. 2009; 8: 3222–38.
КАС пабмед Статья Google Scholar
Дрёзе С., Брандт У., Виттиг И. Комплексы митохондриальной дыхательной цепи как источники и мишени окислительно-восстановительной регуляции на основе тиолов. Биохим Биофиз Акта. 1844; 2014: 1344–54.
Google Scholar
Zarse K, Schmeisser S, Groth M, Priebe S, Beuster G, Kuhlow D, Guthke R, Platzer M, Kahn CR, Ristow M. Нарушение передачи сигналов инсулина-/IGF1 увеличивает продолжительность жизни за счет стимулирования митохондриального пролина катаболизм, вызывающий временный сигнал АФК.Клеточный метаб. 2012;15:451–65.
Нарушение передачи сигналов инсулина-/IGF1 увеличивает продолжительность жизни за счет стимулирования митохондриального пролина катаболизм, вызывающий временный сигнал АФК.Клеточный метаб. 2012;15:451–65.
КАС пабмед ПабМед Центральный Статья Google Scholar
Glasauer A, Chandel NS. РОС. Карр Биол. 2013;23:R100–2.
КАС пабмед Статья Google Scholar
Sena LA, Chandel NS. Физиологические роли митохондриальных активных форм кислорода. Мол Ячейка. 2012;48:158–67.
КАС пабмед ПабМед Центральный Статья Google Scholar
Финкель Т. Передача сигналов митохондриальными оксидантами. Дж. Биол. Хим. 2012; 287:4434–40.
КАС пабмед Статья Google Scholar
Юн Дж., Финкель Т. Митогормезис.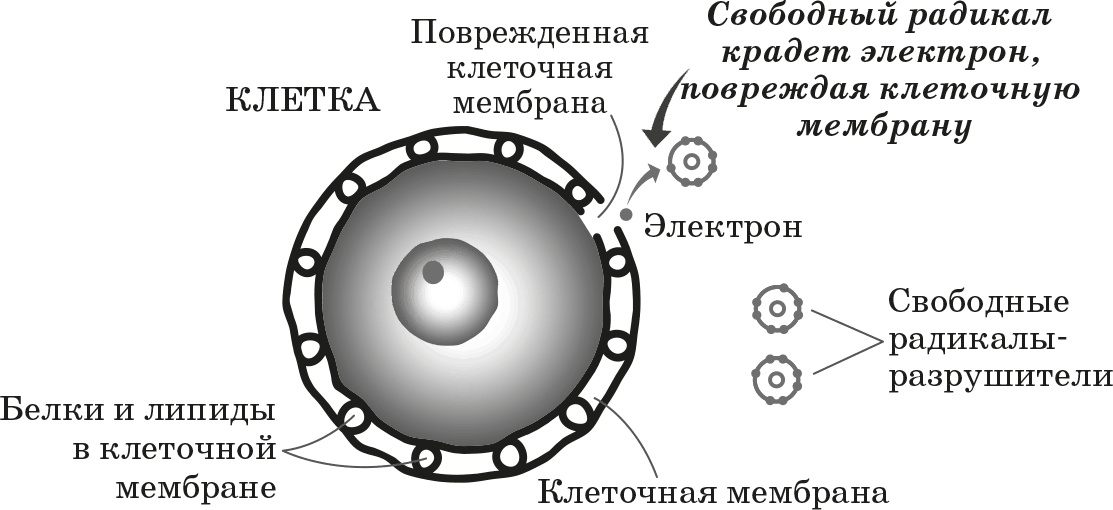 Клеточный метаб. 2014;19:757–66.
Клеточный метаб. 2014;19:757–66.
КАС пабмед ПабМед Центральный Статья Google Scholar
Рахман К. Исследования свободных радикалов, антиоксидантов и сопутствующих факторов.Clin Interv Старение. 2007; 2: 219–36.
КАС пабмед ПабМед Центральный Google Scholar
Molavian H, Tonekaboni AM, Kohandel M, Sivaloganathan S. Синергетическое взаимодействие клеточных антиоксидантов глутатионпероксидазы/пероксиредоксина и других антиоксидантов и его влияние на концентрацию H 2 O 2 . Научный доклад 2015; 5: 1–8.
Артикул Google Scholar
Sheng Y, Abreu IA, Cabelli DE, Maroney MJ, Miller A, Teixeira M, Valentine JS. Супероксиддисмутазы и супероксидредуктазы. Chem Rev. 2014; 114:3854–918.
КАС пабмед ПабМед Центральный Статья Google Scholar

Schmölz L, Birringer M, Lorkowski S, Wallert M. Сложность метаболизма витамина Е. World J Biol Chem. 2016;7:14–43.
ПабМед ПабМед Центральный Статья Google Scholar
Сингх У., Деварадж С., Джалал И. Витамин Е, окислительный стресс и воспаление. Энн Рев Нутр. 2005; 25: 151–74.
КАС Статья Google Scholar
Ван X, Куинн П.Дж. Расположение и функция витамина Е в мембранах (обзор). мол. биол. 2000; 17: 143–56.
ПабМед Статья Google Scholar
Коджо С. Витамин С: основной метаболизм и его функция как показатель окислительного стресса.Курр Мед Хим. 2004; 11:1041–64.
КАС пабмед Статья Google Scholar
Birk J, Meyer M, Aller I, Hansen HG, Odermatt A, Dick TP, Meyer AJ, Appenzeller-Herzog C. Эндоплазматический ретикулум: новый взгляд на восстановленный и окисленный глутатион. Дж. Клеточные науки. 2013; 126:1604–17.
Эндоплазматический ретикулум: новый взгляд на восстановленный и окисленный глутатион. Дж. Клеточные науки. 2013; 126:1604–17.
КАС пабмед Статья Google Scholar
Бхарат С., Хсу М., Каур Д., Раджагопалан С., Андерсен Дж.К.Глутатион, железо и болезнь Паркинсона. Биохим Фармакол. 2002;64:1037–48.
КАС пабмед Статья Google Scholar
Warner DS, Sheng H, Batinic’-Haberle I. Оксиданты, антиоксиданты и ишемический мозг. J Эксперт Биол. 2004; 207:3221–31.
КАС пабмед Статья Google Scholar
Пасторе А., Федеричи Г., Бертини Э., Пьемонте Ф.Анализ глутатиона: участие в окислительно-восстановительном процессе и детоксикации. Клин Чим Акта. 2003; 333:19–39.
КАС пабмед Статья Google Scholar

Mills BJ, Weiss MM, Lang CA, Liu MC, Ziegler C. Изменения глутатиона и цистеина в крови при сердечно-сосудистых заболеваниях. J Lab Clin Med. 2000; 135:396–402.
КАС пабмед Статья Google Scholar
Наварро Х., Обрадор Э., Карретеро Х., Петшен И., Авиньо Х., Перес П., Эстрела Х.М. Изменения статуса глутатиона и антиоксидантной системы в крови и раковых клетках связаны с ростом опухоли in vivo. Свободный Радик Биол Мед. 1999; 26:410–8.
КАС пабмед Статья Google Scholar
Rubartelli A, Bonifaci N, Sitia R. Высокая скорость секреции тиоредоксина коррелирует с остановкой роста клеток гепатомы. Рак рез.1995; 55: 675–80.
КАС пабмед Google Scholar
Tonissen KF, Di Trapani G. Ингибиторы системы тиоредоксин как медиаторы апоптоза для лечения рака. Мол Нутр Фуд Рез. 2009; 53:87–103.
2009; 53:87–103.
КАС пабмед Статья Google Scholar
Yoshida S, Katoh T, Tetsuka T, Uno K, Matsui N, Okamoto T. Участие тиоредоксина в развитии ревматоидного артрита: его костимуляторная роль в TNF-альфа-индуцированной продукции IL-6 и IL-8 из культивированных синовиальных фибробластов.Дж Иммунол. 1999; 163:351-8.
КАС пабмед Google Scholar
Накамура Х., Де Роса С., Редерер М., Андерсон М.Т., Дабс Дж.Г., Йодои Дж., Холмгрен А., Герценберг Л.А., Герценберг Л.А. Повышение уровня тиоредоксина в плазме у ВИЧ-инфицированных. Инт Иммунол. 1996; 8: 603–11.
КАС пабмед Статья Google Scholar
Пакер Л., Рой С., Сен К.К.Альфа-липоевая кислота: метаболический антиоксидант и потенциальный окислительно-восстановительный модулятор транскрипции. Adv Pharmacol. 1997; 38: 79–101.
КАС пабмед Статья Google Scholar
Сеноглу М., Нацитархан В., Курутас Э.Б., Сеноглу Н., Алтун И., Атли Ю., Озбаг Д. Внутрибрюшинная альфа-липоевая кислота для предотвращения повреждения нервной системы после раздавливания седалищного нерва крысы. J Инъекция периферического нерва плечевого сплетения. 2009; 4:22–7.
ПабМед ПабМед Центральный Google Scholar
Belge Kurutas E, Inanc Guler F, Kilinc M. Свободные радикалы. Arch Med Rev J. 2004; 13:120–32.
Google Scholar
Knight TR, Fariss MW, Farhood A, Jaeschke H. Роль перекисного окисления липидов как механизма повреждения печени после передозировки ацетаминофена у мышей. Токсикол науч. 2003; 6: 229–36.
Артикул КАС Google Scholar
Байт М.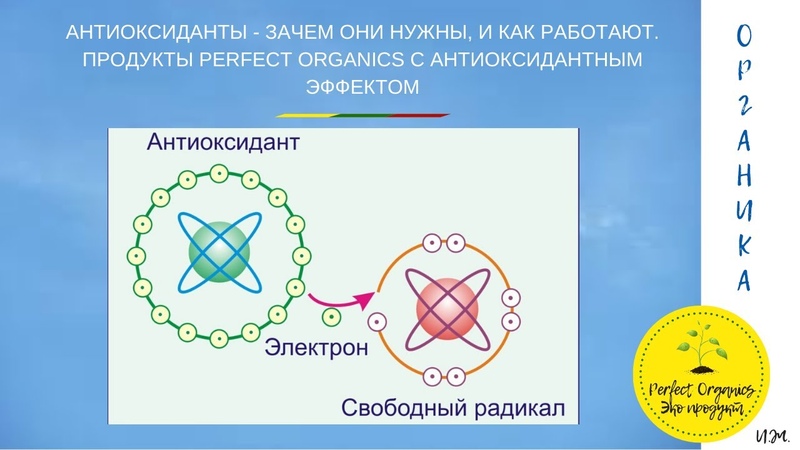 Л., Найт Т.Р., Лемастерс Дж.Дж., Яшке Х.Вызванный ацетаминофеном окислительный стресс и повреждение клеток в культивируемых гепатоцитах мыши: защита с помощью N-ацетилцистеина. Токсикол науч. 2004; 80: 343–9.
Л., Найт Т.Р., Лемастерс Дж.Дж., Яшке Х.Вызванный ацетаминофеном окислительный стресс и повреждение клеток в культивируемых гепатоцитах мыши: защита с помощью N-ацетилцистеина. Токсикол науч. 2004; 80: 343–9.
КАС пабмед Статья Google Scholar
Бренер П.Х., Баллардо М., Мариани Г., Сернадас JMC. Лекарственная ошибка у новорожденного с экстремально низкой массой тела при рождении: передозировка парацетамола. Arch Argent Pediatr. 2013; 111:53–5.
ПабМед Google Scholar
Четинкая А., Бюльбюльоглу Э., Бельге Куруташ Э., Чиралик Х., Кантарчекен Б., Буюкбесе М.А. Благотворное влияние n-ацетилцистеина на колит, вызванный уксусной кислотой, у крыс. Тохоку J Exp Med. 2005; 206: 131–9.
КАС пабмед Статья Google Scholar
Reiter RJ, Acuña-Castroviejo D, Tan DX, Burkhardt S.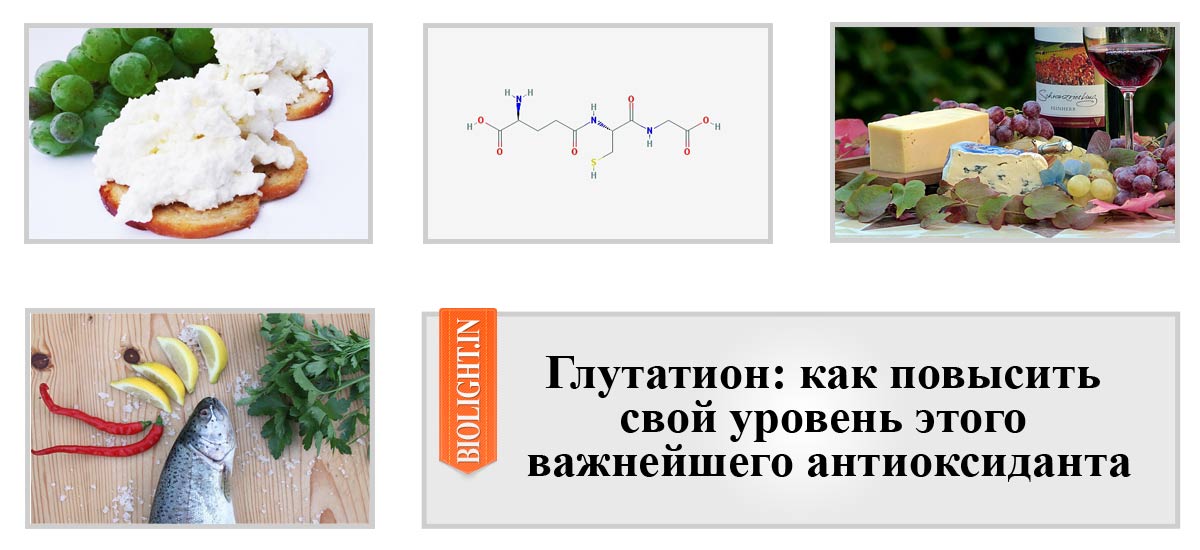 Молекулярное повреждение, опосредованное свободными радикалами. Механизмы защитного действия мелатонина в центральной нервной системе.Энн Н.Ю. Академия наук. 2001; 939: 200–15.
Молекулярное повреждение, опосредованное свободными радикалами. Механизмы защитного действия мелатонина в центральной нервной системе.Энн Н.Ю. Академия наук. 2001; 939: 200–15.
КАС пабмед Статья Google Scholar
Федор Дж., Бурда К. Потенциальная роль каротиноидов как антиоксидантов в здоровье и болезнях человека. Питательные вещества. 2014; 6: 466–88.
ПабМед ПабМед Центральный Статья КАС Google Scholar
Rice-Evans CA, Miller NJ, Paganga G. Взаимосвязь структуры и антиоксидантной активности флавоноидов и фенольных кислот.Свободный Радик Биол Мед. 1996; 20: 933–56.
КАС пабмед Статья Google Scholar
Махима Р.А., Деб Р., Латиф С.К., Абдул Самад Х., Тивари Р., Верма А.К., Кумар А., Дхама К. Иммуномодулирующий и терапевтический потенциал растительных, традиционных/местных и этноветеринарных лекарственных средств. Пак Дж. Биол. Науки. 2012; 15:754–74.
КАС пабмед Статья Google Scholar
Joshipura KJ, Ascherio A, Manson JE, Stampfer MJ, Rimm EB, Speizer FE, Hennekens CH, Spiegelman D, Willett WC. Потребление фруктов и овощей в связи с риском ишемического инсульта. ДЖАМА. 1999; 282:12331–1239.
Артикул Google Scholar
Кокс Б.Д., Уичелоу М.Дж., Прево А.Т. Сезонное потребление салатных овощей и свежих фруктов в связи с развитием сердечно-сосудистых заболеваний и рака. Нутр общественного здравоохранения.2000; 3:19–29.
КАС пабмед Статья Google Scholar
Иствуд, Массачусетс. Взаимодействие пищевых антиоксидантов in vivo: как фрукты и овощи предотвращают болезни? QJM. 1999; 92: 527–30.
КАС пабмед Статья Google Scholar
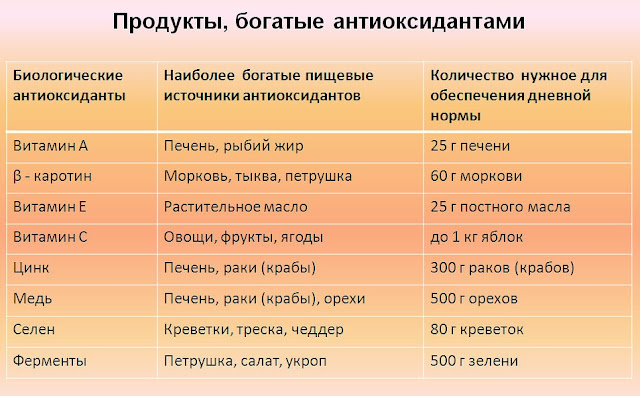
Гилгун-Шерки Ю., Розенбаум З., Меламед Э., Оффен Д. Антиоксидантная терапия при остром поражении центральной нервной системы: современное состояние.Pharmacol Rev. 2002; 54:271–84.
КАС пабмед Статья Google Scholar
Гилгун-Шерки Ю., Меламед Э., Оффен Д. Антиоксидантная терапия при болезни Альцгеймера: современное состояние. Джей Мол Нейроски. 2003; 21:1–11.
КАС пабмед Статья Google Scholar
Алтун И., Курутас Э.Б. Уровни G-белково-связанных рецепторов эстрогена после повреждения периферического нерва на экспериментальной модели крысы.Мировой нейрохирург. 2015; 84: 1903–1906.
ПабМед Статья Google Scholar
Вивеканантхан Д.П., Пенн М.С., Сапп С.К., Хсу А., Тополь Э.Дж. Использование витаминов-антиоксидантов для профилактики сердечно-сосудистых заболеваний: метаанализ рандомизированных исследований. Ланцет. 2003;361:2017–23.
Ланцет. 2003;361:2017–23.
КАС пабмед Статья Google Scholar
Бельг Курутас Э., Сеноглу М., Юксель К.З., Унсал В., Алтун И.Окислительный/нитрозативный стресс у пациентов с модическими изменениями: предварительное контролируемое исследование. Позвоночник (Фила Па, 1976). 2015;40:1101–7.
Артикул Google Scholar
Block KI, Koch AC, Mead MN, Tothy PK, Newman RA, Gyllenhaal C. Влияние антиоксидантных добавок на химиотерапевтическую эффективность: систематический обзор данных рандомизированных контролируемых исследований. Лечение рака, ред. 2007; 33:407–18.
КАС пабмед Статья Google Scholar
Конклин К.А. Окислительный стресс, связанный с химиотерапией: влияние на эффективность химиотерапии. Интегр Рак Тер. 2004; 3: 294–300.
КАС пабмед Статья Google Scholar
Лавенда Б.Д., Келли К.М., Ладас Э.Дж., Сагар С.М., Викерс А., Блумберг Дж.Б. Следует ли избегать дополнительного введения антиоксидантов во время химиотерапии и лучевой терапии? J Natl Cancer Inst. 2008; 100:773–83.
КАС пабмед Статья Google Scholar
Hrelia S, Bordoni A, Angeloni C, Leoncini E, Biagi P. Пищевые вмешательства для противодействия окислительному стрессу в клетках сердца. Ital J Biochem. 2004; 53: 157–63.
КАС пабмед Google Scholar
Ники Э. Антиоксиданты: основные принципы, новые концепции и проблемы. Биомед Дж. 2014; 37:106–11.
ПабМед Google Scholar
Ники Э.Нарушают ли антиоксиданты передачу сигналов активными формами кислорода и продуктами окисления липидов? ФЭБС лат. 2012; 586:3767–70.
КАС пабмед Статья Google Scholar
Рахал А., Кумар А., Сингх В., Ядав Б., Тивари Р., Чакраборти С., Дхама К. Окислительный стресс, прооксиданты и антиоксиданты: взаимодействие. Биомед Рез Инт. 2014; 2014: 1–19.
Артикул КАС Google Scholar
Со М.И., Ли С.М. Защитный эффект низкой дозы аскорбиновой кислоты на гепатобилиарную функцию при ишемии/реперфузии печени у крыс. J Гепатол. 2002; 36: 72–7.
КАС пабмед Статья Google Scholar
Бюттнер Г.Р., Юркевич Б.А. Каталитические металлы, аскорбат и свободные радикалы: комбинации, которых следует избегать. Рад Рез. 1996; 145: 532–41.
КАС Статья Google Scholar
Бюттнер ГР. Порядок расположения свободных радикалов и антиоксидантов: перекисное окисление липидов, α-токоферол и аскорбат. Арх Биохим Биофиз. 1993; 300: 535–43.
КАС пабмед Статья Google Scholar
Яо Х.Т., Луо М.Н., Хунг Л.Б., Чан М.Т., Линь Д.Х., Лии К.К., Хуан К.И. Влияние олигосахаридов хитозана на ферменты, метаболизирующие лекарственные средства, в печени и почках крыс. Пищевая химическая токсикол. 2012;50:1171–7.
КАС пабмед Статья Google Scholar
Hebbar V, Shen G, Hu R, Kim BR, Chen C, Korytko PJ, Crowell JA, Levine BS, Kong AN. Токсикогеномика ресвератрола в печени крыс. Жизнь наук. 2005; 76: 2299–314.
КАС пабмед Статья Google Scholar
Янг А.Дж., Лоу Г.М. Антиоксидантные и прооксидантные свойства каротиноидов. Арх Биохим Биофиз. 2001; 385: 20–7.
КАС пабмед Статья Google Scholar
Контуш А., Финк Б., Картен Б., Кольшюттер А., Бейзигель У. Антиоксидантная и прооксидантная активность α-токоферола в плазме человека и липопротеинов низкой плотности. J липидный рез. 1996; 37: 1436–48.
КАС пабмед Google Scholar
Halliwell B. Являются ли полифенолы антиоксидантами или прооксидантами? Что мы узнаем из клеточных культур и исследований in vivo? Арх Биохим Биофиз. 2008; 476:107–12.
КАС пабмед Статья Google Scholar
Окислители и антиоксиданты | СпрингерЛинк
‘) вар корзинаStepActive = истина var buybox = документ.querySelector(«[data-id=id_»+ метка времени +»]»).parentNode ;[].slice.call(buybox.querySelectorAll(«.вариант-покупки»)).forEach(initCollapsibles) функция initCollapsibles(подписка, индекс) { var toggle = подписка.querySelector(«.цена-варианта-покупки») подписка.classList.remove(«расширенный») var form = подписка.querySelector(«.форма-варианта-покупки») если (форма && cartStepActive) { var formAction = форма.получить атрибут («действие») form.setAttribute(«действие», formAction.replace(«/checkout», «/cart»)) document.querySelector(«#ecommerce-scripts»).addEventListener(«load», bindModal(form, formAction, timestamp, index), false) } var priceInfo = подписка.querySelector(«.Информация о цене») var PurchaseOption = toggle.parentElement если (переключить && форма && priceInfo) { переключать.setAttribute(«роль», «кнопка») toggle.setAttribute(«tabindex», «0») toggle.addEventListener («щелчок», функция (событие) { var expand = toggle.getAttribute(«aria-expanded») === «true» || ложный toggle.setAttribute(«aria-expanded», !expanded) form.hidden = расширенный если (! расширено) { покупкаВариант.classList.add («расширенный») } еще { покупкаOption.classList.remove(«расширенный») } priceInfo.hidden = расширенный }, ложный) } } функция bindModal (форма, formAction, метка времени, индекс) { var weHasBrowserSupport = window.fetch && Array.from функция возврата () { var Buybox = EcommScripts ? EcommScripts.Ящик для покупок: ноль var Modal = EcommScripts ? EcommScripts.Modal : ноль if (weHasBrowserSupport && Buybox && Modal) { var modalID = «ecomm-modal_» + метка времени + «_» + индекс var modal = новый модальный (modalID) modal.domEl.addEventListener («закрыть», закрыть) функция закрыть () { форма.querySelector(«кнопка[тип=отправить]»).фокус() } форма.setAttribute( «действие», formAction.replace(«/checkout», «/cart?messageOnly=1») ) form.addEventListener( «Отправить», Buybox.interceptFormSubmit( Буйбокс.fetchFormAction(окно.fetch), Buybox.triggerModalAfterAddToCartSuccess(модальный), консоль.лог, ), ложный ) document.body.appendChild(modal.domEl) } } } функция initKeyControls() { документ.addEventListener(«keydown», функция (событие) { if (document.activeElement.classList.contains(«цена-варианта-покупки») && (event.code === «Пробел» || event.code === «Enter»)) { если (document.activeElement) { событие.preventDefault() документ.activeElement.click() } } }, ложный) } функция InitialStateOpen() { var buyboxWidth = buybox.смещениеШирина ;[].slice.call(buybox.querySelectorAll(«.опция покупки»)).forEach(функция (опция, индекс) { var toggle = option.querySelector(«.цена-варианта-покупки») var form = option.querySelector(«.форма-варианта-покупки») var priceInfo = option.querySelector(«.Информация о цене») если (buyboxWidth > 480) { переключить.щелчок() } еще { если (индекс === 0) { переключать.щелчок() } еще { toggle.setAttribute («ария-расширенная», «ложь») form.hidden = «скрытый» priceInfo.hidden = «скрытый» } } }) } начальное состояниеОткрыть() если (window.buyboxInitialized) вернуть window.buyboxInitialized = истина initKeyControls() })()Антиоксидант — обзор
Применение антиоксидантов в косметике
Антиоксиданты отвечают за разрыв цепи поглотителей радикалов и за ингибирование реакции окисления; с помощью этих механизмов антиоксиданты могут предотвращать окислительное повреждение [56,57].В косметических препаратах антиоксиданты выполняют две функции: как активные ингредиенты и как защитники других ингредиентов от окисления [38].
В настоящее время применение антиоксидантов в косметике увеличивается; однако, чтобы получить желаемую активность, следует рассмотреть некоторые стратегии. Короткое время жизни АФК можно преодолеть, используя антиоксиданты, обладающие высокой реакционной способностью и емкостью. Антиоксиданты не должны превращаться в свои радикалы, такие как аскорбил- или токофериловые радикалы; это вызовет цепную реакцию.Антиоксиданты должны оставаться стабильными в продукте; они не должны реагировать с другими ингредиентами и должны быть защищены от радикалов кислорода. Выбор антиоксидантов, которые можно использовать в косметике, зависит от их гидрофобных или липофильных характеристик. К сожалению, иногда выбор антиоксиданта(ов) (фармацевтической промышленностью), используемого в косметических продуктах, основывается не на научных суждениях, а скорее на их цене.
Как правило, антиоксиданты по своей природе нестабильны, сильно окрашены и подвержены гидролизу и фотодеградации в присутствии кислорода; вот почему очень трудно иметь хорошие косметические рецептуры и поддерживать их эстетическую ценность и приемлемость.Модификация химической структуры антиоксиданта, например замена его эфирами (например, токоферилацетатом, аскорбилпальмитатом) или укорочение липофильной цепи CoQ10, может улучшить его стабильность, но, к сожалению, снижает его активность.
Чтобы быть активным, необходим стабильный антиоксидант, но, к сожалению, антиоксиданты, как правило, являются нестабильными соединениями. Эта нестабильность может вызвать много проблем. В косметической рецептуре концентрация антиоксидантов должна быть стабильной для достижения желаемой активности.Их цвет не должен изменяться в процессе производства и хранения, чтобы их антиоксидантная активность оставалась постоянной, а продукт сохранял эстетический вид. Все это порождает множество проблем при рецептуре косметических средств [12,21,32]. Вот почему необходим валидный метод определения способности антиоксиданта оценивать его активность [32].
Применение относительно новой технологии «системы доставки на основе липидов» может защищать и поддерживать стабильность антиоксидантов.Эта технология также обладает защитным действием против обезвоживания кожи. Липидные носители могут увеличить проникновение антиоксиданта в кожу, поэтому его желаемая активность может быть гарантирована. Были разработаны и используются различные липидные носители, такие как эмульсия наночастиц, различные везикулярные системы (липосомы, фитосомы, трансферсомы, этосомы, ниосомы и нанотопы) и системы частиц (липидные микрочастицы и липидные наночастицы). Стабильность аскорбилпальмитата и витаминов К и А в косметике можно повысить, используя липидные наноносители.Фитосомы зеленого чая и виноградных косточек могут улучшить их активность по удалению свободных радикалов и защите от ультрафиолета. Проникновение ацетата витамина Е в кожу было увеличено при использовании Nanotop™.